

Adaptive evolution of Moniliophthora PR-1 proteins towards its pathogenic lifestyle
- PMID: 33990179
- PMCID: PMC8120714
- DOI: 10.1186/s12862-021-01818-5
Adaptive evolution of Moniliophthora PR-1 proteins towards its pathogenic lifestyle
Abstract
Background: Plant pathogenesis related-1 (PR-1) proteins belong to the CAP superfamily and have been characterized as markers of induced defense against pathogens. Moniliophthora perniciosa and Moniliophthora roreri are hemibiotrophic fungi that respectively cause the witches' broom disease and frosty pod rot in Theobroma cacao. Interestingly, a large number of plant PR-1-like genes are present in the genomes of both species and many are up-regulated during the biotrophic interaction. In this study, we investigated the evolution of PR-1 proteins from 22 genomes of Moniliophthora isolates and 16 other Agaricales species, performing genomic investigation, phylogenetic reconstruction, positive selection search and gene expression analysis.
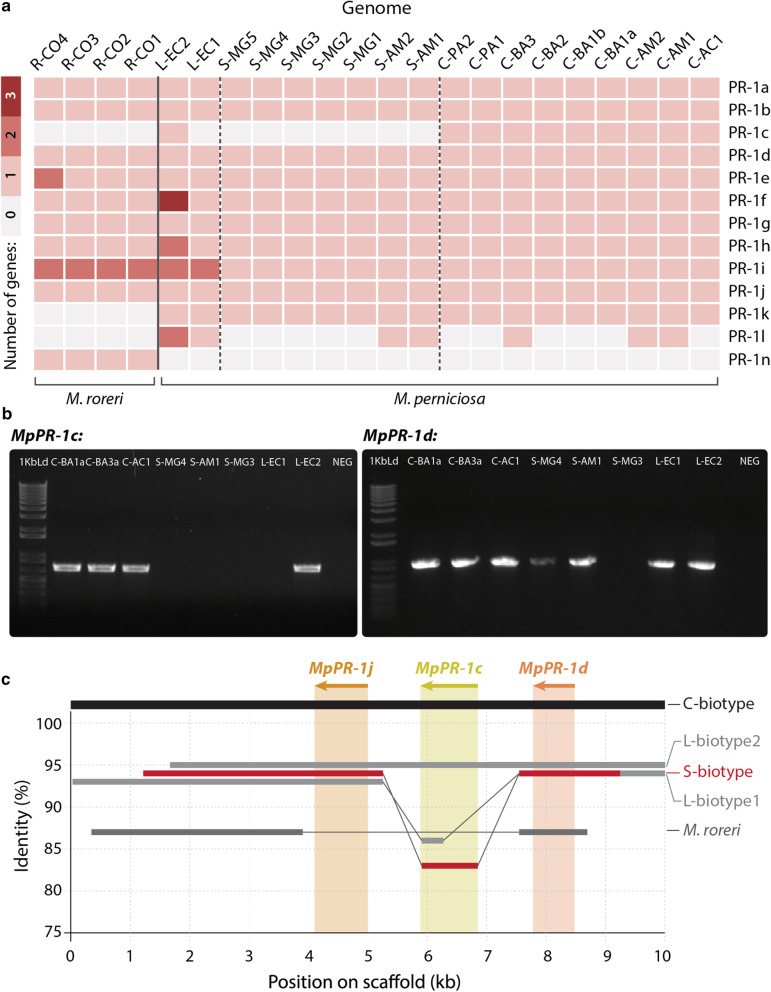
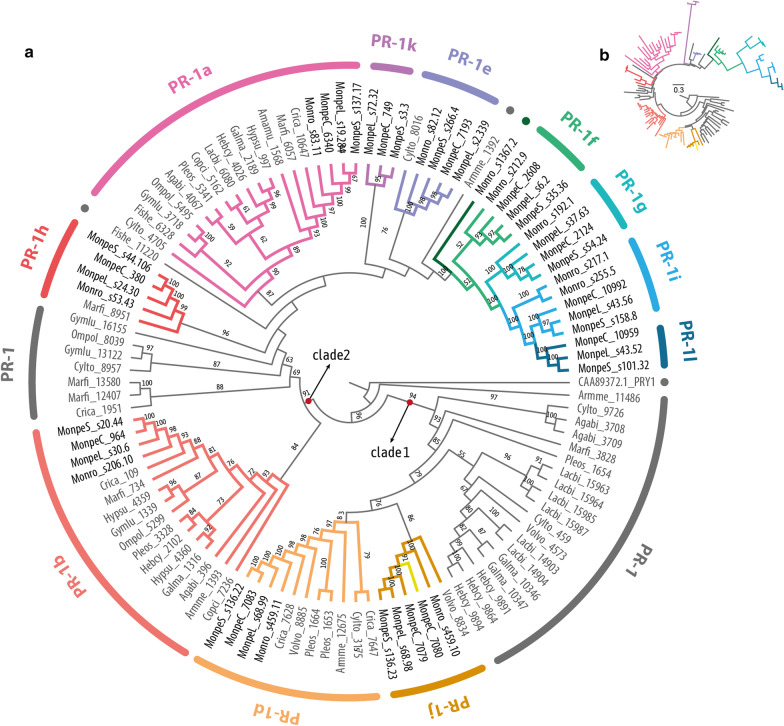
Results: Phylogenetic analysis revealed conserved PR-1 genes (PR-1a, b, d, j), shared by many Agaricales saprotrophic species, that have diversified in new PR-1 genes putatively related to pathogenicity in Moniliophthora (PR-1f, g, h, i), as well as in recent specialization cases within M. perniciosa biotypes (PR-1c, k, l) and M. roreri (PR-1n). PR-1 families in Moniliophthora with higher evolutionary rates exhibit induced expression in the biotrophic interaction and positive selection clues, supporting the hypothesis that these proteins accumulated adaptive changes in response to host-pathogen arms race. Furthermore, although previous work showed that MpPR-1 can detoxify plant antifungal compounds in yeast, we found that in the presence of eugenol M. perniciosa differentially expresses only MpPR-1e, k, d, of which two are not linked to pathogenicity, suggesting that detoxification might not be the main function of most MpPR-1.
Conclusions: Based on analyses of genomic and expression data, we provided evidence that the evolution of PR-1 in Moniliophthora was adaptive and potentially related to the emergence of the parasitic lifestyle in this genus. Additionally, we also discuss how fungal PR-1 proteins could have adapted from basal conserved functions to possible roles in fungal pathogenesis.
Keywords: Adaptation; Fungi; Gene evolution; Phylogenetics; Phytopathogen; Positive selection; Witches’ broom disease.
Conflict of interest statement
The authors declare no competing interests.
Figures




Similar articles
-
Genome sequence and effectorome of Moniliophthora perniciosa and Moniliophthora roreri subpopulations.BMC Genomics. 2018 Jul 3;19(1):509. doi: 10.1186/s12864-018-4875-7. BMC Genomics. 2018. PMID: 29969982 Free PMC article.
-
Moniliophthora roreri, causal agent of cacao frosty pod rot.Mol Plant Pathol. 2018 Jul;19(7):1580-1594. doi: 10.1111/mpp.12648. Epub 2018 Feb 15. Mol Plant Pathol. 2018. PMID: 29194910 Free PMC article.
-
The mitochondrial genome of Moniliophthora roreri, the frosty pod rot pathogen of cacao.Fungal Biol. 2012 May;116(5):551-62. doi: 10.1016/j.funbio.2012.01.008. Epub 2012 Feb 15. Fungal Biol. 2012. PMID: 22559916
-
Moniliophthora perniciosa, the causal agent of witches' broom disease of cacao: what's new from this old foe?Mol Plant Pathol. 2008 Sep;9(5):577-88. doi: 10.1111/j.1364-3703.2008.00496.x. Mol Plant Pathol. 2008. PMID: 19018989 Free PMC article. Review.
-
Chocolate Under Threat from Old and New Cacao Diseases.Phytopathology. 2019 Aug;109(8):1331-1343. doi: 10.1094/PHYTO-12-18-0477-RVW. Epub 2019 Jul 1. Phytopathology. 2019. PMID: 31115251 Review.
Cited by
-
Clonal reproduction of Moniliophthora roreri and the emergence of unique lineages with distinct genomes during range expansion.G3 (Bethesda). 2023 Aug 30;13(9):jkad125. doi: 10.1093/g3journal/jkad125. G3 (Bethesda). 2023. PMID: 37337677 Free PMC article.
References
-
- Asojo OA, Goud G, Dhar K, Loukas A, Zhan B, Deumic V, et al. X-ray structure of Na-ASP-2, a pathogenesis-related-1 protein from the nematode parasite, Necator americanus, and a vaccine antigen for human hookworm infection. J Mol Biol. 2005;346(3):801–814. doi: 10.1016/j.jmb.2004.12.023. - DOI - PubMed
-
- Chalmers IW, McArdle AJ, Coulson RM, Wagner MA, Schmid R, Hirai H, et al. Developmentally regulated expression, alternative splicing and distinct sub-groupings in members of the Schistosomamansoni venom allergen-like (SmVAL) gene family. BMC Genomics. 2008;9(1):89. doi: 10.1186/1471-2164-9-89. - DOI - PMC - PubMed
Publication types
MeSH terms
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous