

NMDAR-dependent long-term depression is associated with increased short term plasticity through autophagy mediated loss of PSD-95
- PMID: 33990590
- PMCID: PMC8121912
- DOI: 10.1038/s41467-021-23133-9
NMDAR-dependent long-term depression is associated with increased short term plasticity through autophagy mediated loss of PSD-95
Abstract
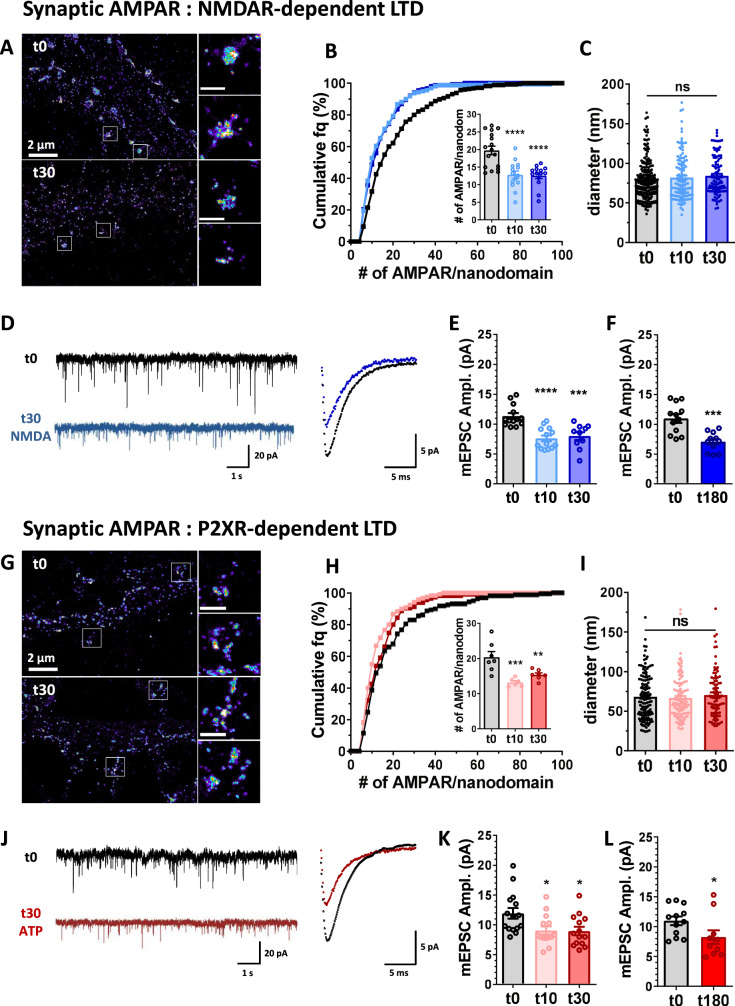
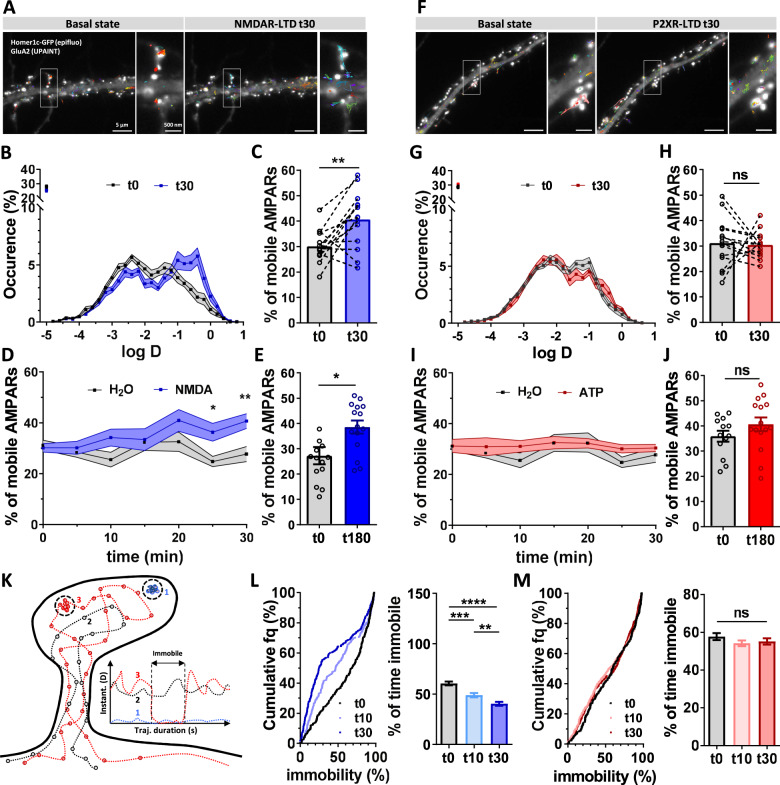
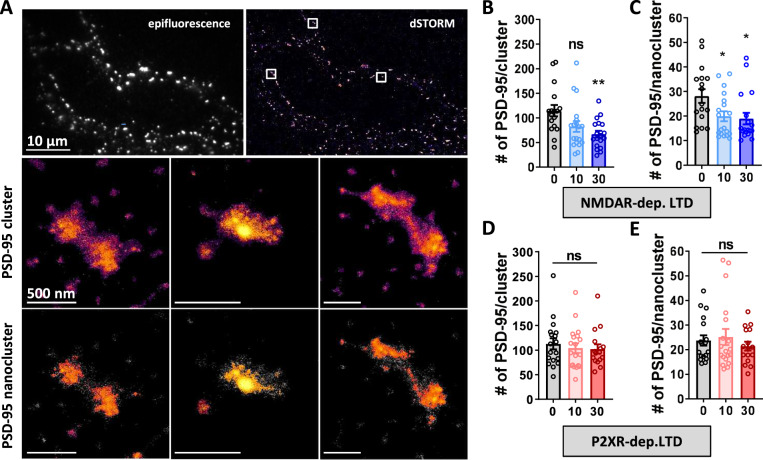
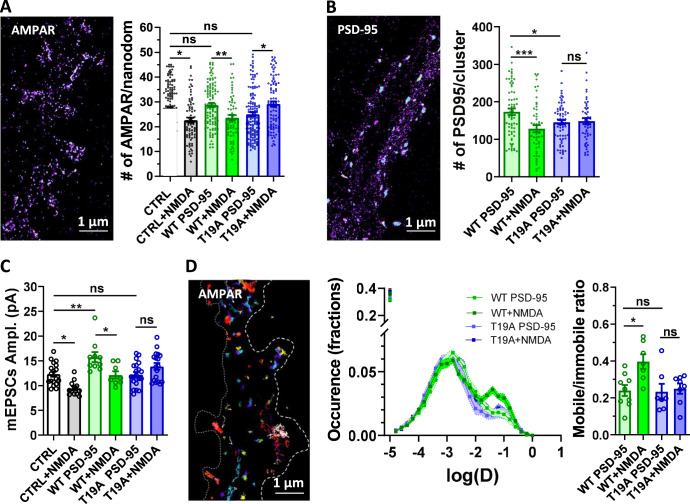
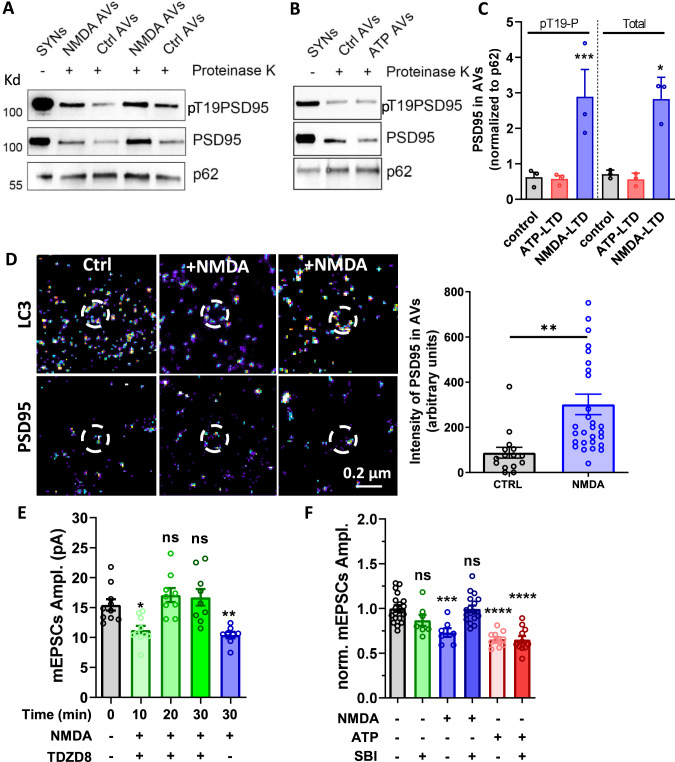
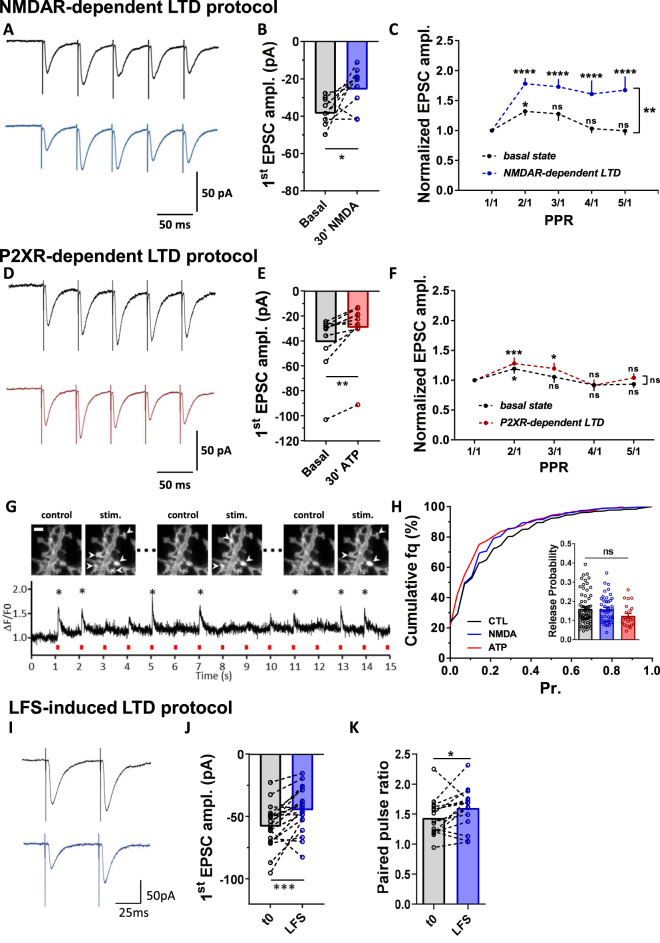
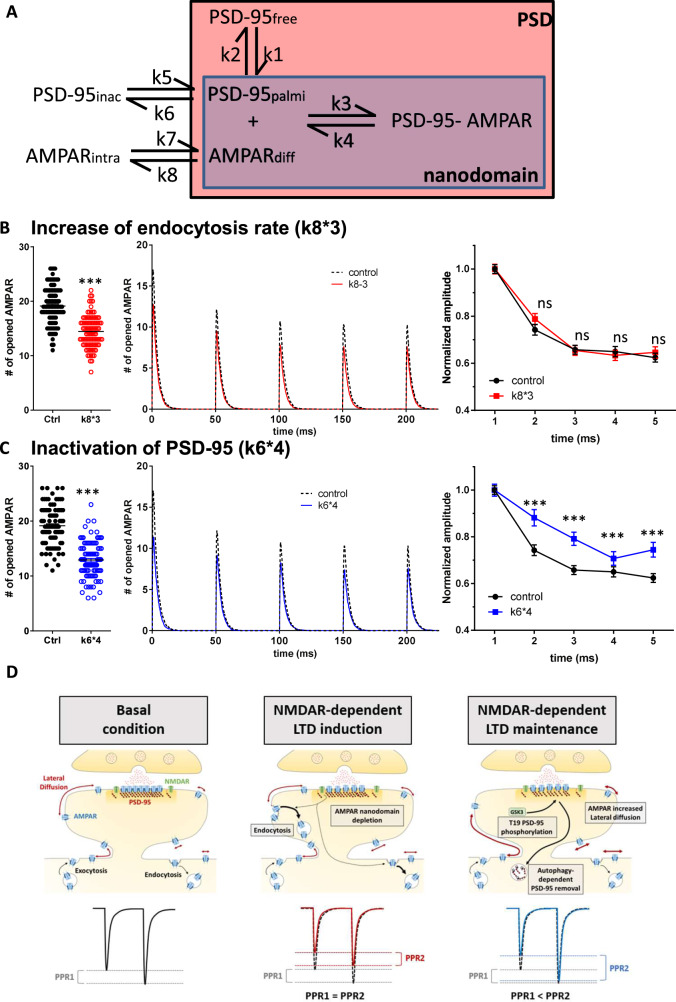
Long-term depression (LTD) of synaptic strength can take multiple forms and contribute to circuit remodeling, memory encoding or erasure. The generic term LTD encompasses various induction pathways, including activation of NMDA, mGlu or P2X receptors. However, the associated specific molecular mechanisms and effects on synaptic physiology are still unclear. We here compare how NMDAR- or P2XR-dependent LTD affect synaptic nanoscale organization and function in rodents. While both LTDs are associated with a loss and reorganization of synaptic AMPARs, only NMDAR-dependent LTD induction triggers a profound reorganization of PSD-95. This modification, which requires the autophagy machinery to remove the T19-phosphorylated form of PSD-95 from synapses, leads to an increase in AMPAR surface mobility. We demonstrate that these post-synaptic changes that occur specifically during NMDAR-dependent LTD result in an increased short-term plasticity improving neuronal responsiveness of depressed synapses. Our results establish that P2XR- and NMDAR-mediated LTD are associated to functionally distinct forms of LTD.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
A critical role for PSD-95/AKAP interactions in endocytosis of synaptic AMPA receptors.Nat Neurosci. 2009 Feb;12(2):172-81. doi: 10.1038/nn.2249. Epub 2009 Jan 25. Nat Neurosci. 2009. PMID: 19169250 Free PMC article.
-
Autophagy controls the induction and developmental decline of NMDAR-LTD through endocytic recycling.Nat Commun. 2020 Jun 12;11(1):2979. doi: 10.1038/s41467-020-16794-5. Nat Commun. 2020. PMID: 32532981 Free PMC article.
-
Regulated RalBP1 binding to RalA and PSD-95 controls AMPA receptor endocytosis and LTD.PLoS Biol. 2009 Sep;7(9):e1000187. doi: 10.1371/journal.pbio.1000187. Epub 2009 Sep 8. PLoS Biol. 2009. PMID: 19823667 Free PMC article.
-
Regulation of neuronal PKA signaling through AKAP targeting dynamics.Eur J Cell Biol. 2006 Jul;85(7):627-33. doi: 10.1016/j.ejcb.2006.01.010. Epub 2006 Feb 28. Eur J Cell Biol. 2006. PMID: 16504338 Review.
-
Hippocampal long-term synaptic plasticity and signal amplification of NMDA receptors.Crit Rev Neurobiol. 2006;18(1-2):71-84. doi: 10.1615/critrevneurobiol.v18.i1-2.80. Crit Rev Neurobiol. 2006. PMID: 17725510 Review.
Cited by
-
Synapse-specific structural plasticity that protects and refines local circuits during LTP and LTD.Philos Trans R Soc Lond B Biol Sci. 2024 Jul 29;379(1906):20230224. doi: 10.1098/rstb.2023.0224. Epub 2024 Jun 10. Philos Trans R Soc Lond B Biol Sci. 2024. PMID: 38853547 Free PMC article.
-
The needs of a synapse-How local organelles serve synaptic proteostasis.EMBO J. 2022 Apr 4;41(7):e110057. doi: 10.15252/embj.2021110057. Epub 2022 Mar 14. EMBO J. 2022. PMID: 35285533 Free PMC article. Review.
-
Glutamate delta 1 receptor regulates autophagy mechanisms and affects excitatory synapse maturation in the somatosensory cortex.Pharmacol Res. 2022 Apr;178:106144. doi: 10.1016/j.phrs.2022.106144. Epub 2022 Mar 15. Pharmacol Res. 2022. PMID: 35304260 Free PMC article.
-
GlyT1 Inhibition by NFPS Promotes Neuroprotection in Amyloid-β-Induced Alzheimer's Disease Animal Model.Neurochem Res. 2024 Sep;49(9):2535-2555. doi: 10.1007/s11064-024-04190-0. Epub 2024 Jun 18. Neurochem Res. 2024. PMID: 38888830
-
Modulation of autophagy by melatonin and its receptors: implications in brain disorders.Acta Pharmacol Sin. 2025 Mar;46(3):525-538. doi: 10.1038/s41401-024-01398-2. Epub 2024 Oct 24. Acta Pharmacol Sin. 2025. PMID: 39448859 Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
