

The principles of directed cell migration
- PMID: 33990789
- PMCID: PMC8663916
- DOI: 10.1038/s41580-021-00366-6
The principles of directed cell migration
Abstract
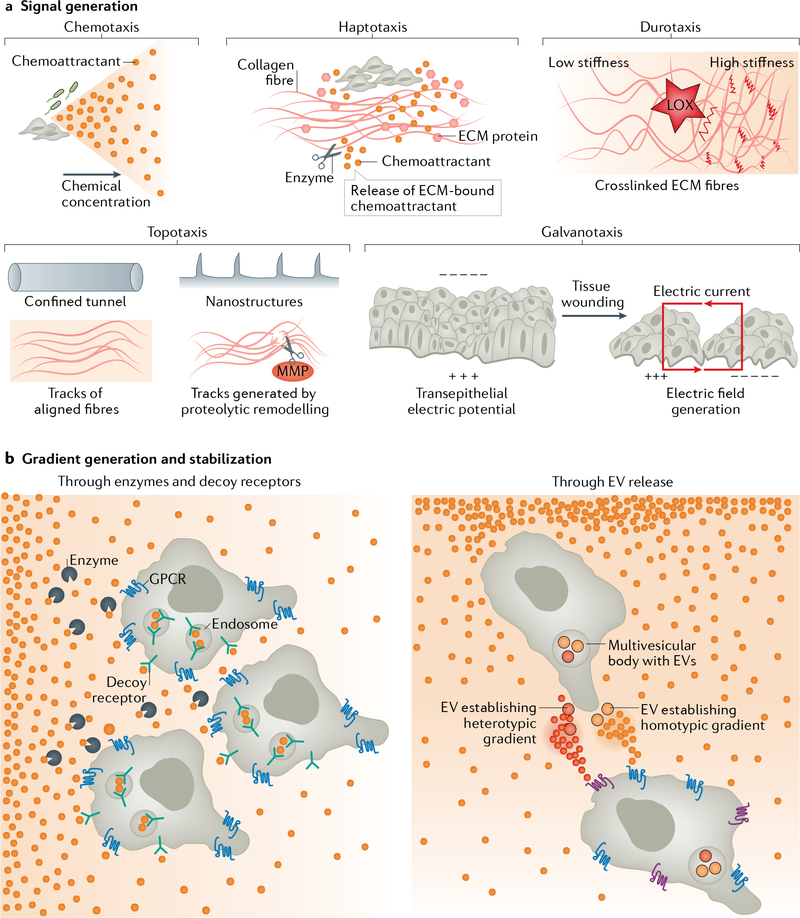
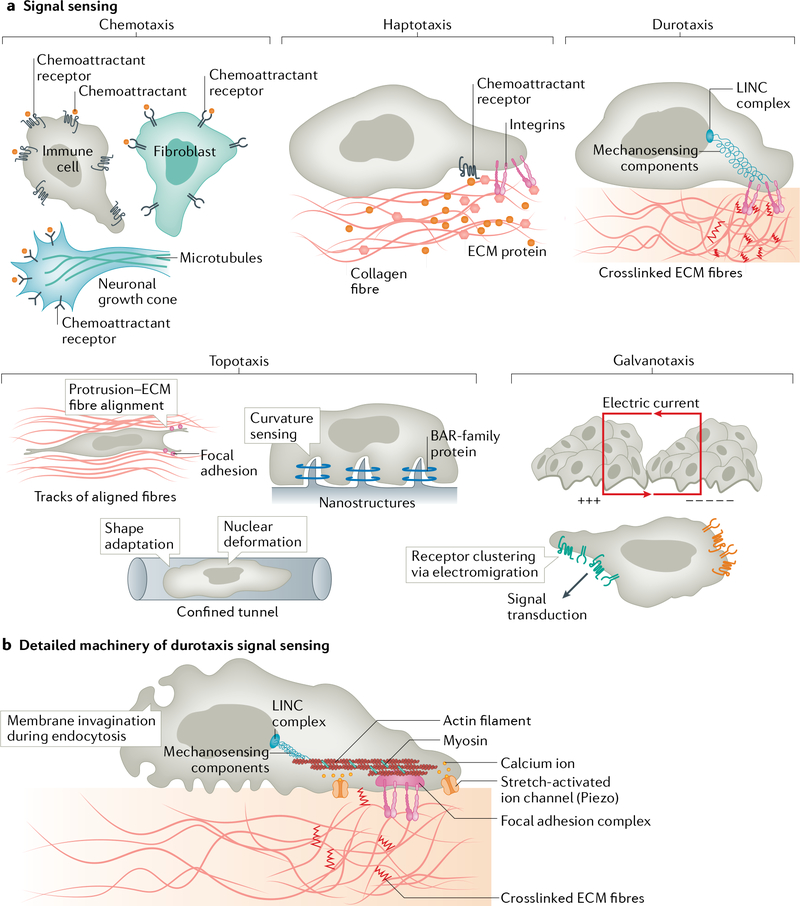
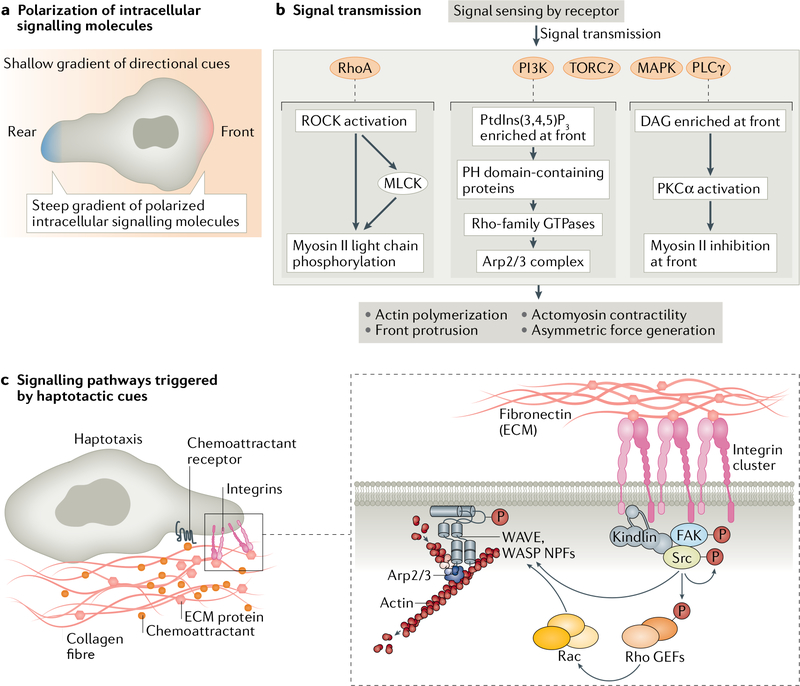
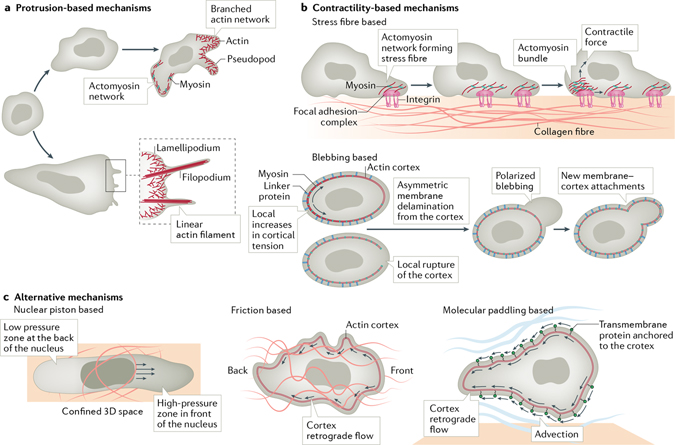
Cells have the ability to respond to various types of environmental cues, and in many cases these cues induce directed cell migration towards or away from these signals. How cells sense these cues and how they transmit that information to the cytoskeletal machinery governing cell translocation is one of the oldest and most challenging problems in biology. Chemotaxis, or migration towards diffusible chemical cues, has been studied for more than a century, but information is just now beginning to emerge about how cells respond to other cues, such as substrate-associated cues during haptotaxis (chemical cues on the surface), durotaxis (mechanical substrate compliance) and topotaxis (geometric features of substrate). Here we propose four common principles, or pillars, that underlie all forms of directed migration. First, a signal must be generated, a process that in physiological environments is much more nuanced than early studies suggested. Second, the signal must be sensed, sometimes by cell surface receptors, but also in ways that are not entirely clear, such as in the case of mechanical cues. Third, the signal has to be transmitted from the sensing modules to the machinery that executes the actual movement, a step that often requires amplification. Fourth, the signal has to be converted into the application of asymmetric force relative to the substrate, which involves mostly the cytoskeleton, but perhaps other players as well. Use of these four pillars has allowed us to compare some of the similarities between different types of directed migration, but also to highlight the remarkable diversity in the mechanisms that cells use to respond to different cues provided by their environment.
© 2021. Springer Nature Limited.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
References
-
- Janssen E & Geha RS Primary immunodeficiencies caused by mutations in actin regulatory proteins. Immunol. Rev. 287, 121–134 (2019). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
