

Parallel Rap1>RalGEF>Ral and Ras signals sculpt the C. elegans nervous system
- PMID: 33991533
- PMCID: PMC8277711
- DOI: 10.1016/j.ydbio.2021.05.004
Parallel Rap1>RalGEF>Ral and Ras signals sculpt the C. elegans nervous system
Abstract
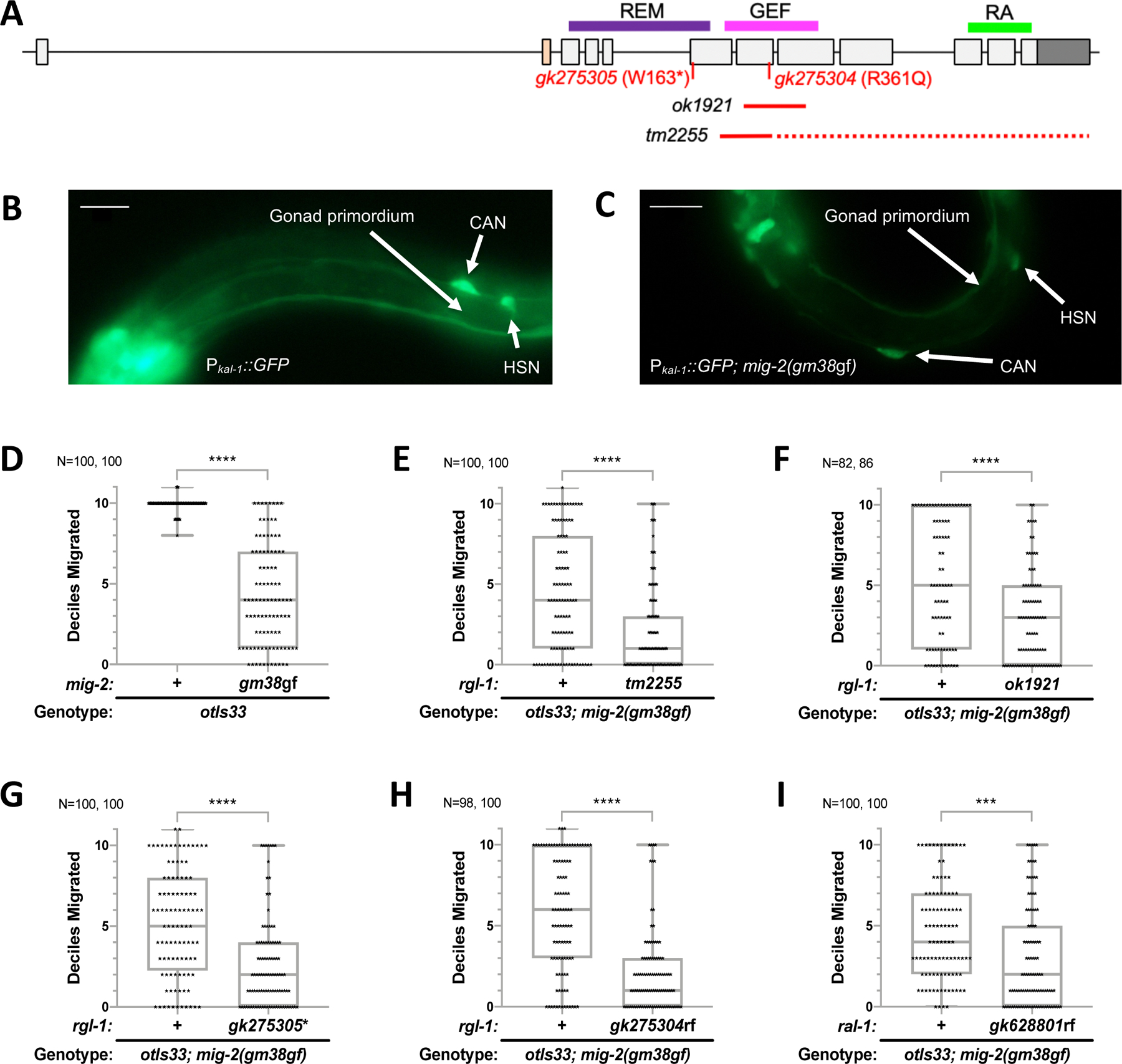
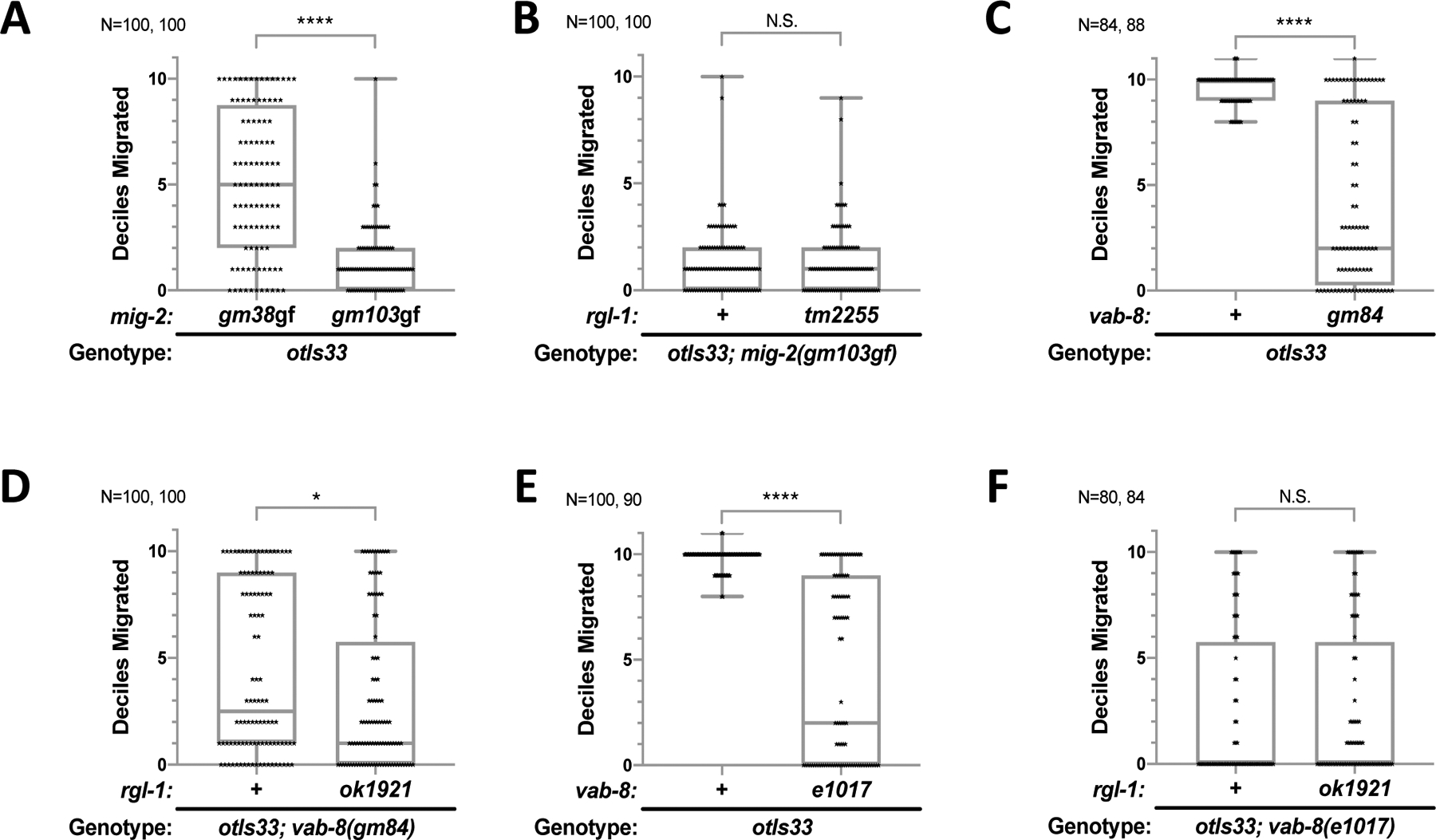
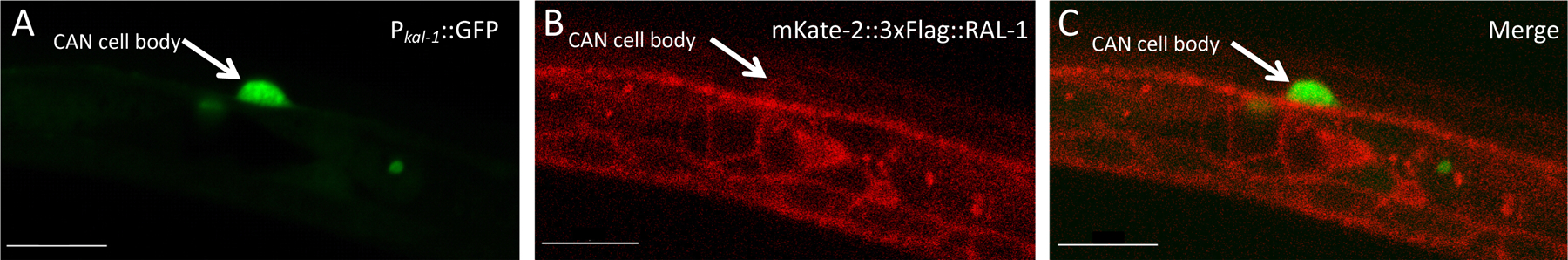
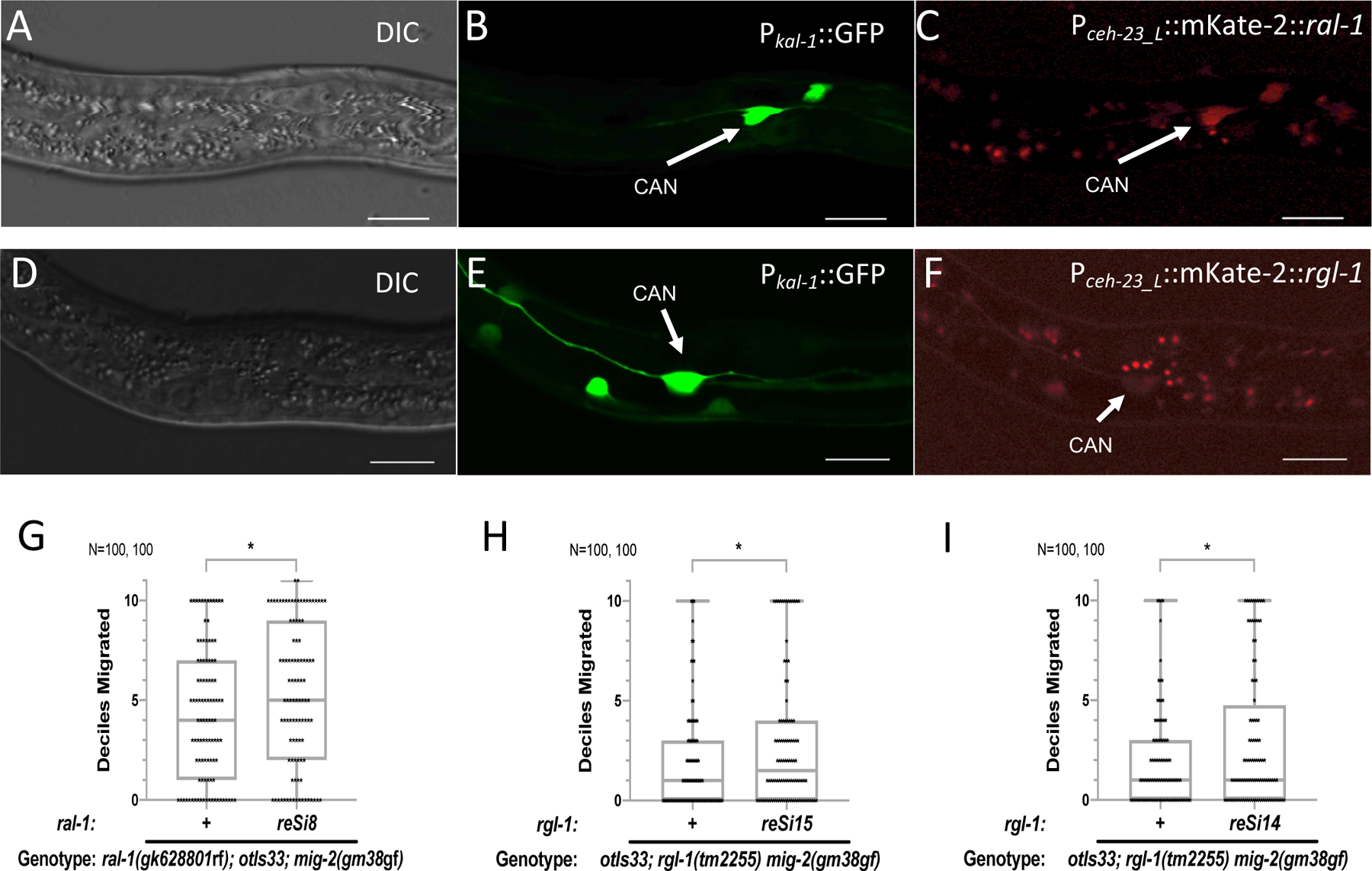
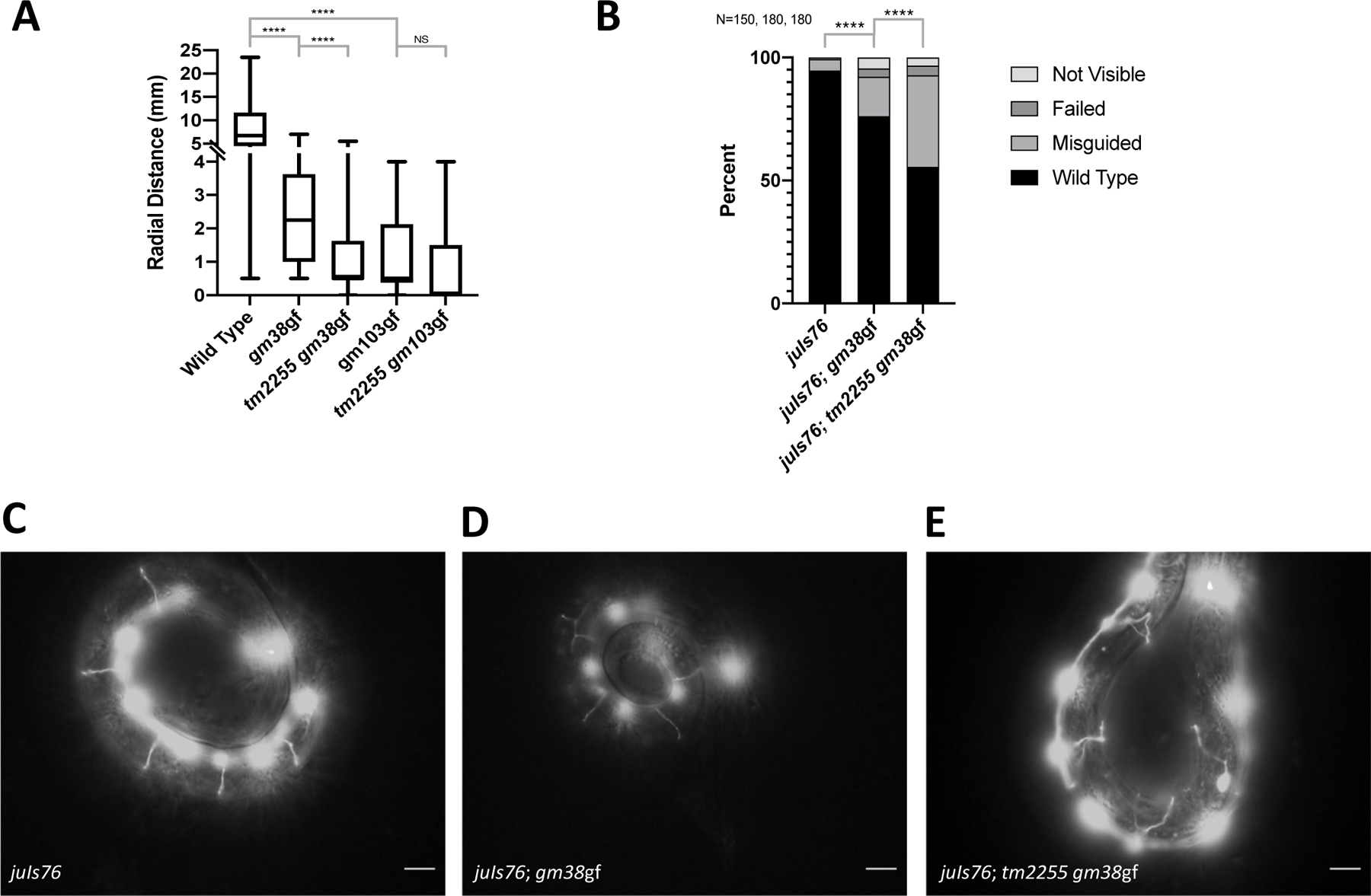
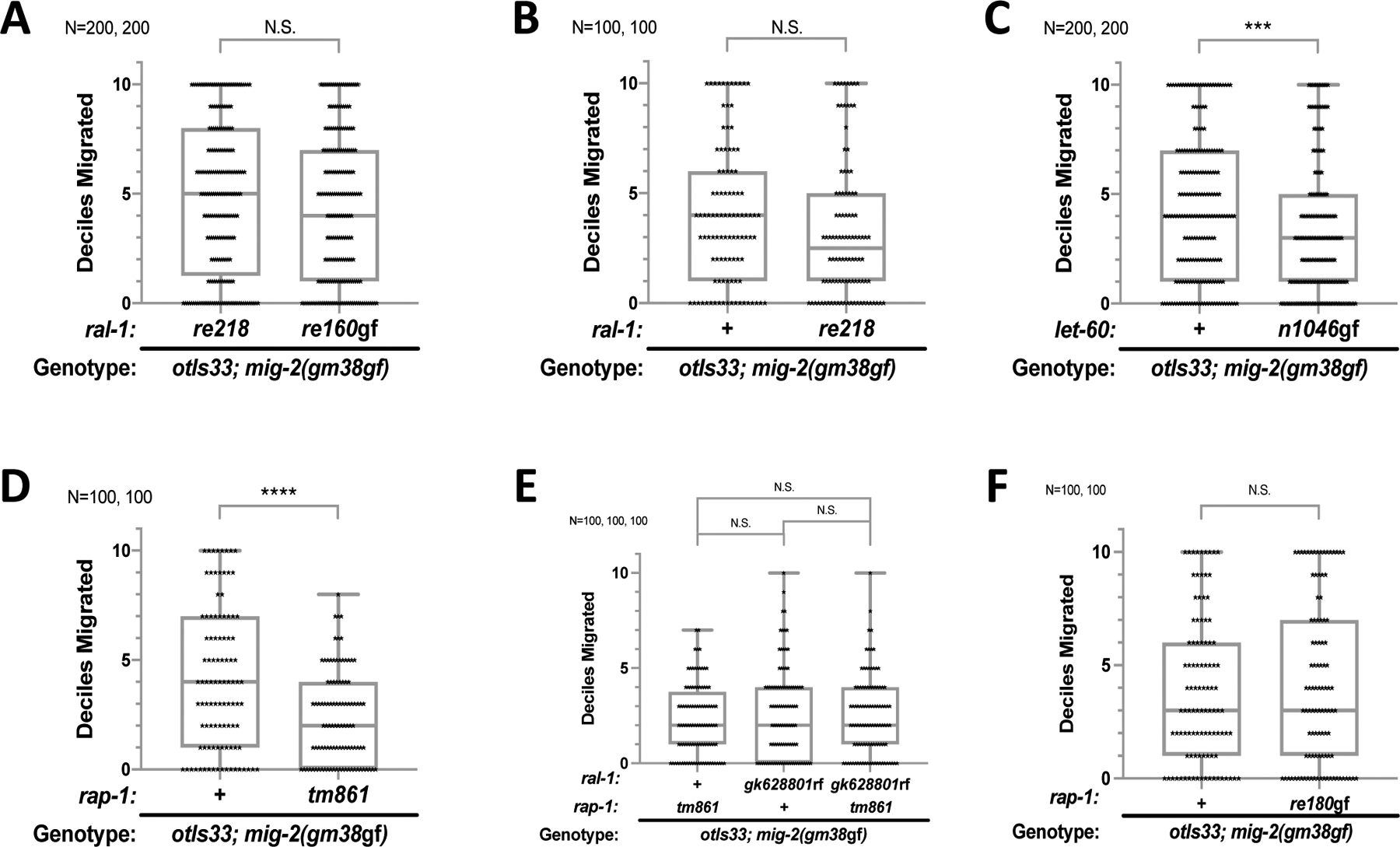
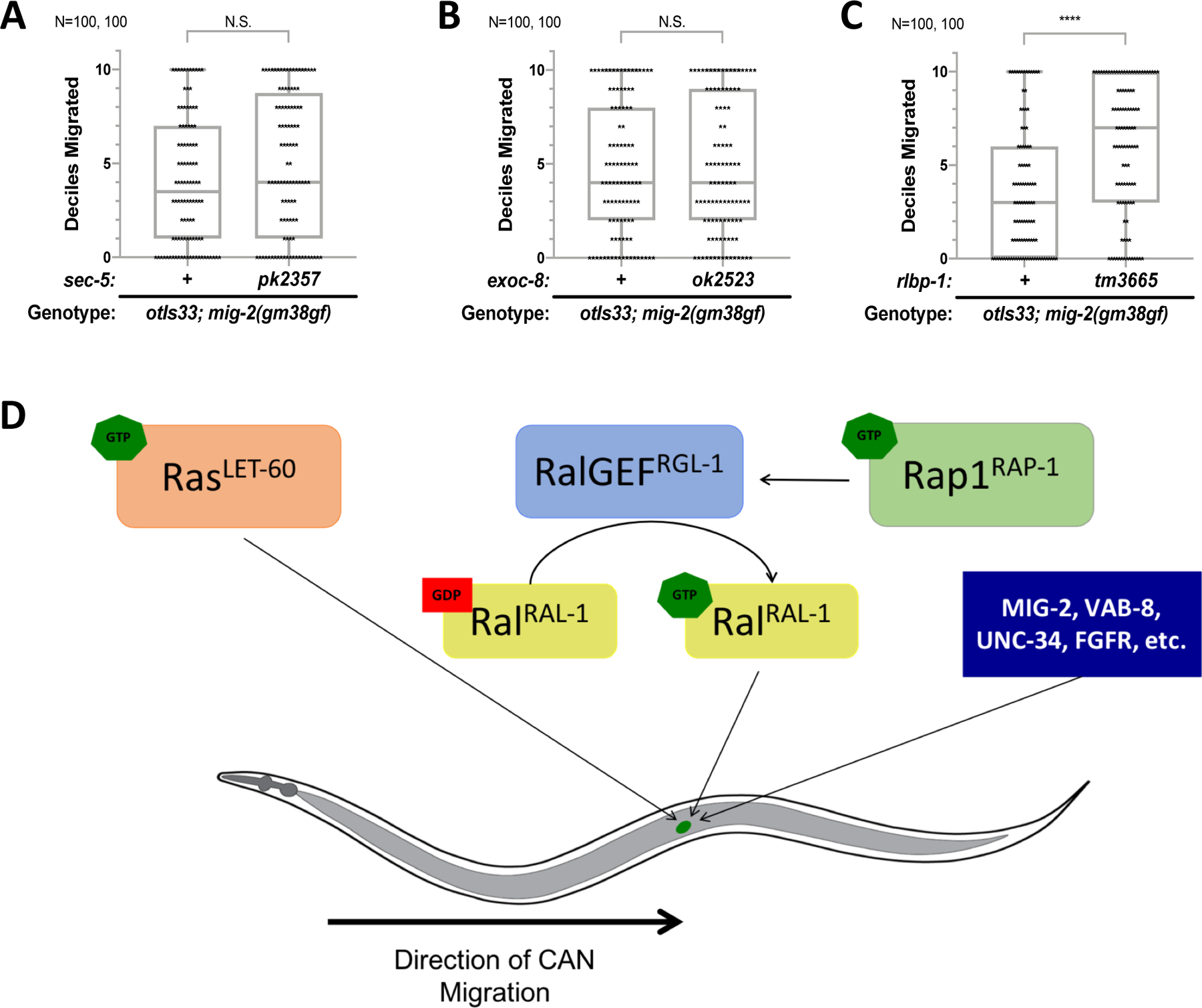
Ras is the most commonly mutated oncogene in humans and uses three oncogenic effectors: Raf, PI3K, and RalGEF activation of Ral. Understanding the importance of RalGEF>Ral signaling in cancer is hampered by the paucity of knowledge about their function in animal development, particularly in cell movements. We found that mutations that disrupt function of RalGEF or Ral enhance migration phenotypes of mutants for genes with established roles in cell migration. We used as a model the migration of the canal associated neurons (CANs), and validated our results in HSN cell migration, neurite guidance, and general animal locomotion. These functions of RalGEF and Ral are specific to their control of Ral signaling output rather than other published functions of these proteins. In this capacity Ral functions cell autonomously as a permissive developmental signal. In contrast, we observed Ras, the canonical activator of RalGEF>Ral signaling in cancer, to function as an instructive signal. Furthermore, we unexpectedly identified a function for the close Ras relative, Rap1, consistent with activation of RalGEF>Ral. These studies define functions of RalGEF>Ral, Rap1 and Ras signaling in morphogenetic processes that fashion the nervous system. We have also defined a model for studying how small GTPases partner with downstream effectors. Taken together, this analysis defines novel molecules and relationships in signaling networks that control cell movements during development of the nervous system.
Keywords: Exo84; Exocyst; LET-60; LIN-45; RAL-1; RAP-1; RGL-1; RalBP1; Sec5.
Copyright © 2021 Elsevier Inc. All rights reserved.
Figures
References
-
- Awasthi S, Cheng JZ, Singhal SS, Pandya U, Sharma R, Singh SV, Zimniak P, and Awasthi YC (2001). Functional reassembly of ATP-dependent xenobiotic transport by the N- and C-terminal domains of RLIP76 and identification of ATP binding sequences. Biochemistry 40, 4159–4168. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
