

Cross-Whisker Adaptation of Neurons in Layer 2/3 of the Rat Barrel Cortex
- PMID: 33994963
- PMCID: PMC8113387
- DOI: 10.3389/fnsys.2021.646563
Cross-Whisker Adaptation of Neurons in Layer 2/3 of the Rat Barrel Cortex
Abstract
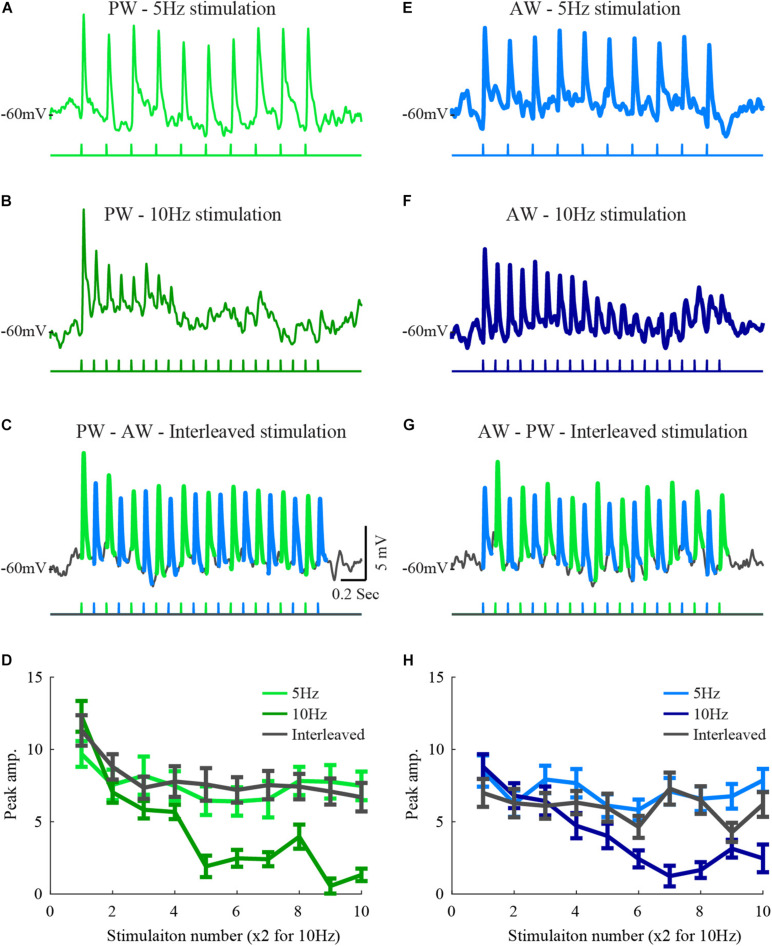
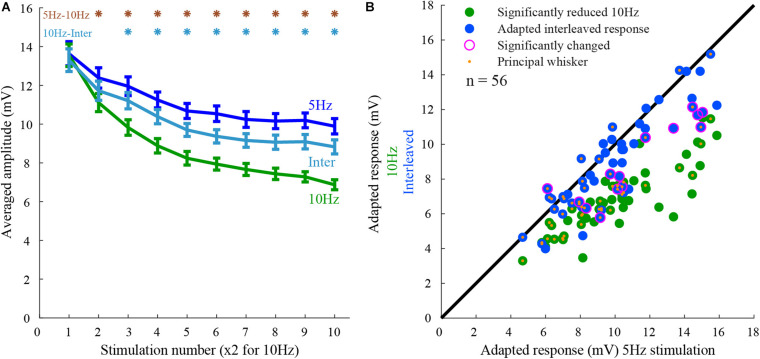
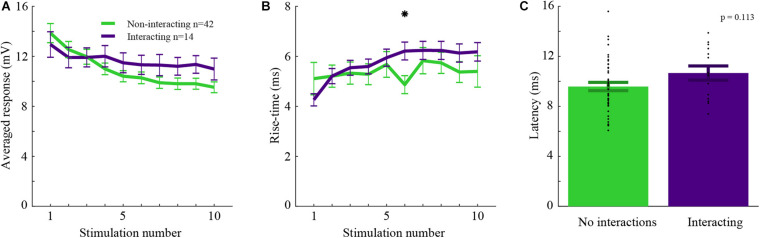
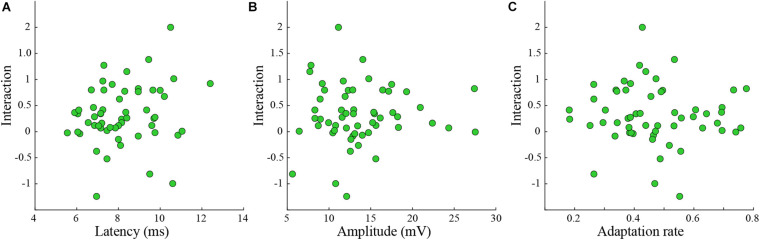
Neurons in the barrel cortex respond preferentially to stimulation of one principal whisker and weakly to several adjacent whiskers. Such integration exists already in layer 4, the pivotal recipient layer of thalamic inputs. Previous studies show that cortical neurons gradually adapt to repeated whisker stimulations and that layer 4 neurons exhibit whisker specific adaptation and no apparent interactions with other whiskers. This study aimed to study the specificity of adaptation of layer 2/3 cortical cells. Towards this aim, we compared the synaptic response of neurons to either repetitive stimulation of one of two responsive whiskers or when repetitive stimulation of the two whiskers was interleaved. We found that in most layer 2/3 cells adaptation is whisker-specific. These findings indicate that despite the multi-whisker receptive fields in the cortex, the adaptation process for each whisker-pathway is mostly independent of other whiskers. A mechanism allowing high responsiveness in complex environments.
Keywords: barrel cortex; in vivo; integration; intracellular; layer 2/3 cortex; receptive fields; whisker stimulation.
Copyright © 2021 Katz and Lampl.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Armstrong-James M., Callahan C. A. (1991). Thalamo-cortical processing of vibrissal information in the rat. II. spatiotemporal convergence in the thalamic ventroposterior medial nucleus (VPm) and its relevance to generation of receptive fields of S1 cortical “barrel” neurones. J. Comp. Neurol. 303 211–224. 10.1002/cne.903030204 - DOI - PubMed
-
- Armstrong-James M., Callahan C. A., Friedman M. A. (1991). Thalamo-cortical processing of vibrissal information in the rat. I. Intracortical origins of surround but not centre-receptive fields of layer IV neurones in the rat S1 barrel field cortex. J. Comp. Neurol. 303 193–210. 10.1002/cne.903030203 - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources