

Coupling of Transcription and Translation in Archaea: Cues From the Bacterial World
- PMID: 33995325
- PMCID: PMC8116511
- DOI: 10.3389/fmicb.2021.661827
Coupling of Transcription and Translation in Archaea: Cues From the Bacterial World
Abstract
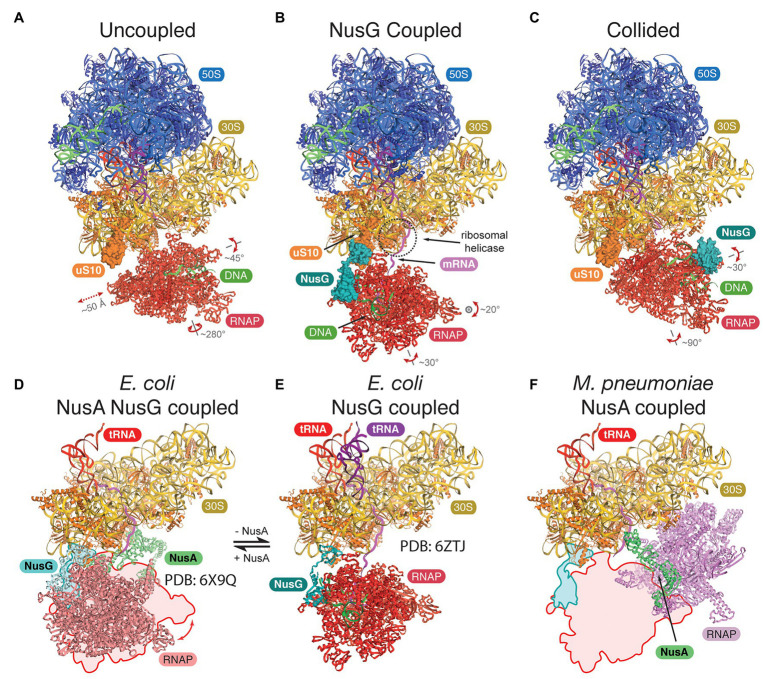
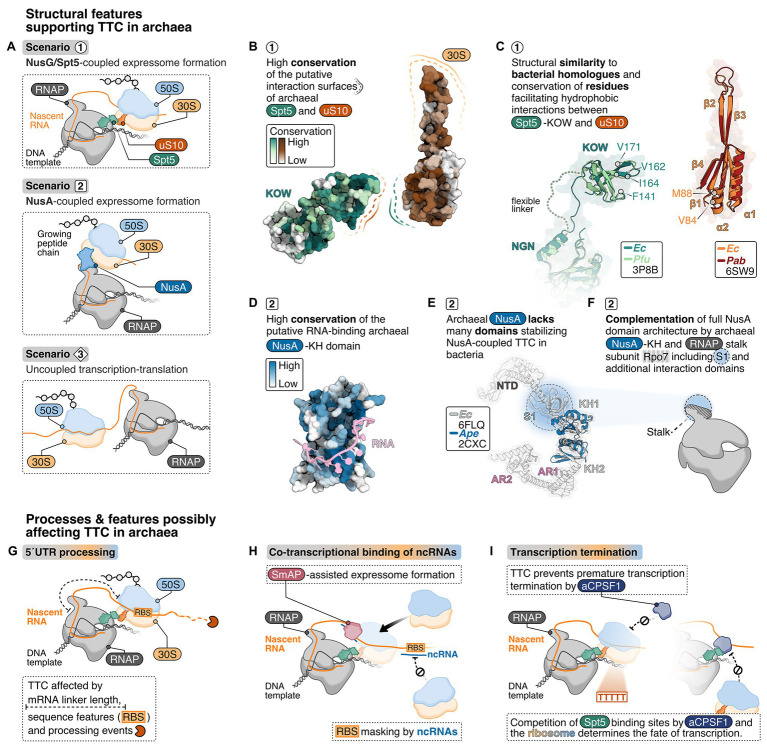
The lack of a nucleus is the defining cellular feature of bacteria and archaea. Consequently, transcription and translation are occurring in the same compartment, proceed simultaneously and likely in a coupled fashion. Recent cryo-electron microscopy (cryo-EM) and tomography data, also combined with crosslinking-mass spectrometry experiments, have uncovered detailed structural features of the coupling between a transcribing bacterial RNA polymerase (RNAP) and the trailing translating ribosome in Escherichia coli and Mycoplasma pneumoniae. Formation of this supercomplex, called expressome, is mediated by physical interactions between the RNAP-bound transcription elongation factors NusG and/or NusA and the ribosomal proteins including uS10. Based on the structural conservation of the RNAP core enzyme, the ribosome, and the universally conserved elongation factors Spt5 (NusG) and NusA, we discuss requirements and functional implications of transcription-translation coupling in archaea. We furthermore consider additional RNA-mediated and co-transcriptional processes that potentially influence expressome formation in archaea.
Keywords: Nus; NusG; RNA polymerase; Spt4/5; archaea; expressome; ribosome.
Copyright © 2021 Weixlbaumer, Grünberger, Werner and Grohmann.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
In-cell architecture of an actively transcribing-translating expressome.Science. 2020 Jul 31;369(6503):554-557. doi: 10.1126/science.abb3758. Science. 2020. PMID: 32732422 Free PMC article.
-
Structural basis of transcription-translation coupling.Science. 2020 Sep 11;369(6509):1359-1365. doi: 10.1126/science.abb5317. Epub 2020 Aug 20. Science. 2020. PMID: 32820061 Free PMC article.
-
Architecture of a transcribing-translating expressome.Science. 2017 Apr 14;356(6334):194-197. doi: 10.1126/science.aal3059. Science. 2017. PMID: 28408604 Free PMC article.
-
Macromolecular assemblies supporting transcription-translation coupling.Transcription. 2021 Aug;12(4):103-125. doi: 10.1080/21541264.2021.1981713. Epub 2021 Sep 27. Transcription. 2021. PMID: 34570660 Free PMC article. Review.
-
NusG/Spt5: are there common functions of this ubiquitous transcription elongation factor?Curr Opin Microbiol. 2014 Apr;18:68-71. doi: 10.1016/j.mib.2014.02.005. Epub 2014 Mar 12. Curr Opin Microbiol. 2014. PMID: 24632072 Free PMC article. Review.
Cited by
-
Bacterial Rps3 counters oxidative and UV stress by recognizing and processing AP-sites on mRNA via a novel mechanism.Nucleic Acids Res. 2024 Dec 11;52(22):13996-14012. doi: 10.1093/nar/gkae1130. Nucleic Acids Res. 2024. PMID: 39588766 Free PMC article.
-
Structural basis of archaeal RNA polymerase transcription elongation and Spt4/5 recruitment.Nucleic Acids Res. 2024 Jun 10;52(10):6017-6035. doi: 10.1093/nar/gkae282. Nucleic Acids Res. 2024. PMID: 38709902 Free PMC article.
-
Co-transcriptional gene regulation in eukaryotes and prokaryotes.Nat Rev Mol Cell Biol. 2024 Jul;25(7):534-554. doi: 10.1038/s41580-024-00706-2. Epub 2024 Mar 20. Nat Rev Mol Cell Biol. 2024. PMID: 38509203 Free PMC article. Review.
-
Chromosome architecture in an archaeal species naturally lacking structural maintenance of chromosomes proteins.Nat Microbiol. 2024 Jan;9(1):263-273. doi: 10.1038/s41564-023-01540-6. Epub 2023 Dec 18. Nat Microbiol. 2024. PMID: 38110698 Free PMC article.
-
The structure and activities of the archaeal transcription termination factor Eta detail vulnerabilities of the transcription elongation complex.Proc Natl Acad Sci U S A. 2022 Aug 9;119(32):e2207581119. doi: 10.1073/pnas.2207581119. Epub 2022 Aug 2. Proc Natl Acad Sci U S A. 2022. PMID: 35917344 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources