

Structure and evolutionary trace-assisted screening of a residue swapping the substrate ambiguity and chiral specificity in an esterase
- PMID: 33995922
- PMCID: PMC8105184
- DOI: 10.1016/j.csbj.2021.04.041
Structure and evolutionary trace-assisted screening of a residue swapping the substrate ambiguity and chiral specificity in an esterase
Abstract
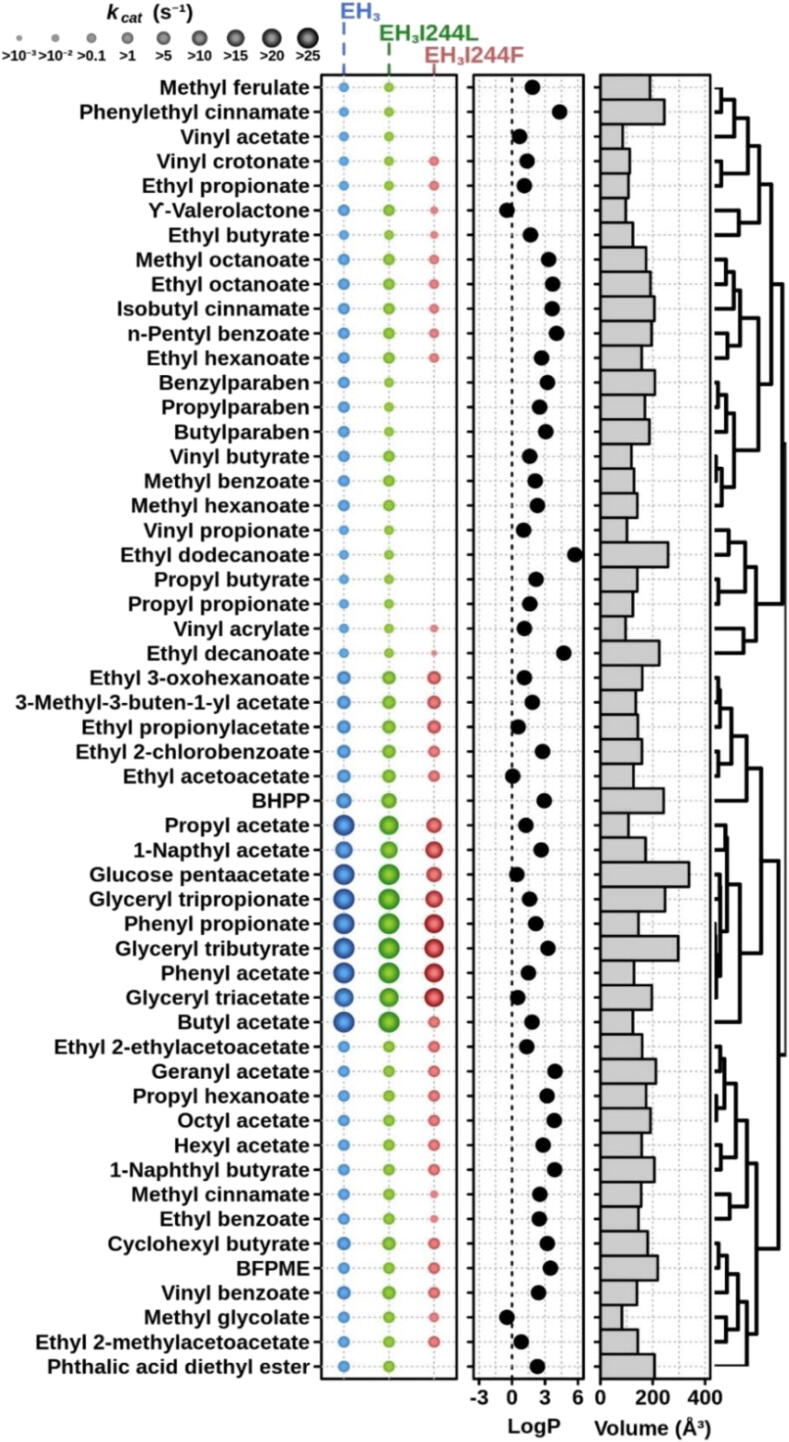
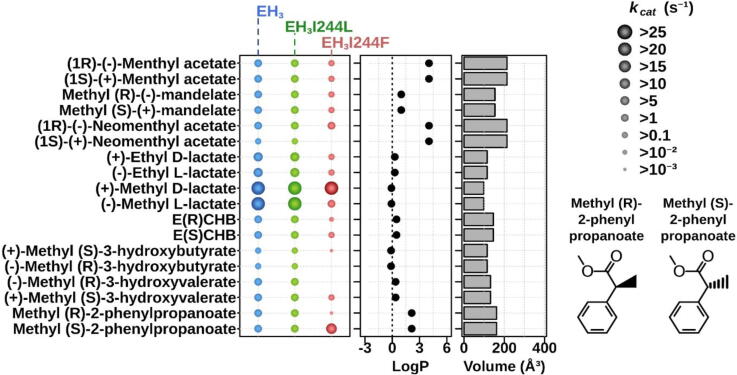
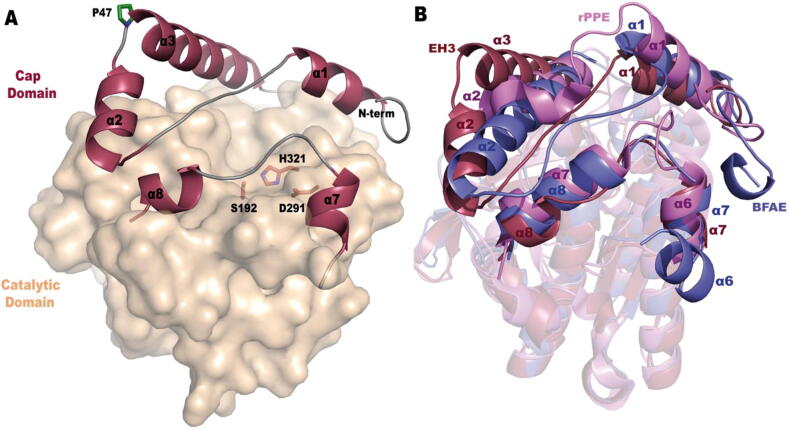
Our understanding of enzymes with high substrate ambiguity remains limited because their large active sites allow substrate docking freedom to an extent that seems incompatible with stereospecificity. One possibility is that some of these enzymes evolved a set of evolutionarily fitted sequence positions that stringently allow switching substrate ambiguity and chiral specificity. To explore this hypothesis, we targeted for mutation a serine ester hydrolase (EH3) that exhibits an impressive 71-substrate repertoire but is not stereospecific (e.e. 50%). We used structural actions and the computational evolutionary trace method to explore specificity-swapping sequence positions and hypothesized that position I244 was critical. Driven by evolutionary action analysis, this position was substituted to leucine, which together with isoleucine appears to be the amino acid most commonly present in the closest homologous sequences (max. identity, ca. 67.1%), and to phenylalanine, which appears in distant homologues. While the I244L mutation did not have any functional consequences, the I244F mutation allowed the esterase to maintain a remarkable 53-substrate range while gaining stereospecificity properties (e.e. 99.99%). These data support the possibility that some enzymes evolve sequence positions that control the substrate scope and stereospecificity. Such residues, which can be evolutionarily screened, may serve as starting points for further designing substrate-ambiguous, yet chiral-specific, enzymes that are greatly appreciated in biotechnology and synthetic chemistry.
Keywords: Crystal structure; EA, evolutionary action; ET, evolutionary trace; Eapp, apparent enantioselectivity; Esterase; Evolutionary trace; HEPES, 40 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid; Ni-NTA, nickel-nitrilotriacetic acid; Promiscuity; Protein engineering; Specificity.
© 2021 The Author(s).
Conflict of interest statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures




References
-
- Jegannathan K.R., Nielsen P.H. Environmental assessment of enzyme use in industrial production – a literature review. J Clean Prod. 2013;42:228–240.
-
- Sheldon R.A., Woodley J.M. Role of biocatalysis in sustainable chemistry. Chem Rev. 2018;118(2):801–838. - PubMed
-
- Tawfik D.S., Gruic-Sovulj I. How evolution shapes enzyme selectivity - lessons from aminoacyl-tRNA synthetases and other amino acid utilizing enzymes. FEBS J. 2020;287:1284–1305. - PubMed
-
- Coscolín C., Martínez-Martínez M., Chow J., Bargiela R., García-Moyano A., Bjerga G. Relationships between substrate promiscuity and chiral selectivity of esterases from phylogenetically and environmentally diverse microorganisms. Catalysts. 2018;8(1):10.
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources