

Development and Characterization of High-Throughput Caenorhabditis elegans - Enterococcus faecium Infection Model
- PMID: 33996637
- PMCID: PMC8116795
- DOI: 10.3389/fcimb.2021.667327
Development and Characterization of High-Throughput Caenorhabditis elegans - Enterococcus faecium Infection Model
Abstract
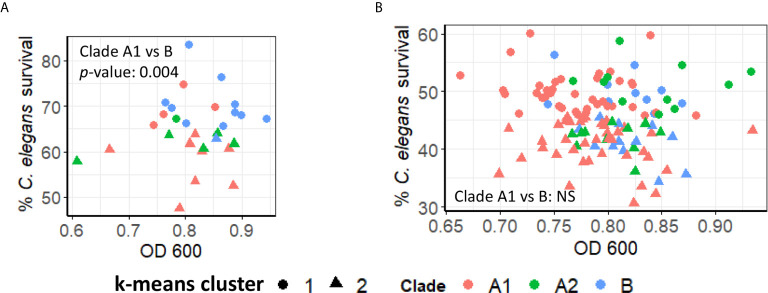
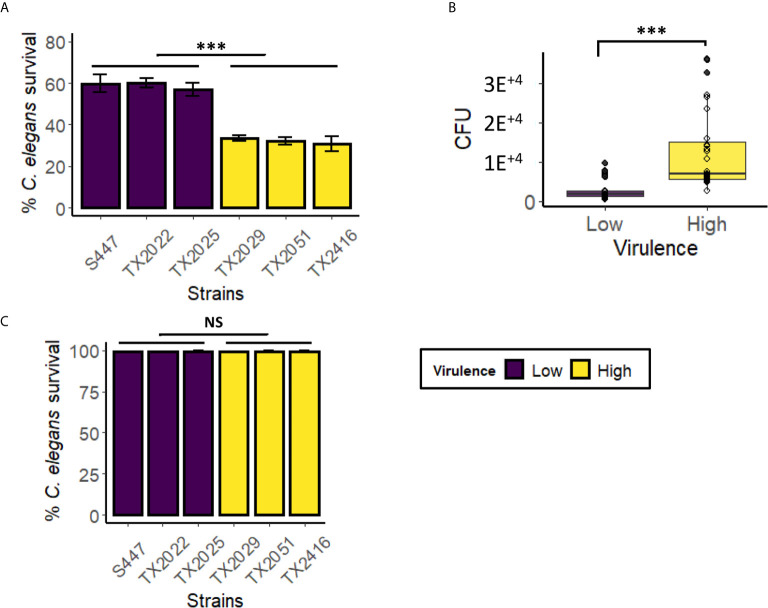
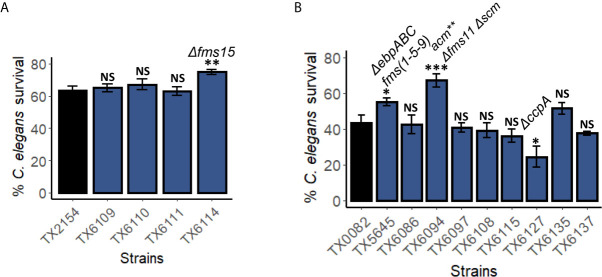
The genus Enterococcus includes two Gram-positive pathogens of particular clinical relevance: E. faecalis and E. faecium. Infections with each of these pathogens are becoming more frequent, particularly in the case of hospital-acquired infections. Like most other bacterial species of clinical importance, antimicrobial resistance (and, specifically, multi-drug resistance) is an increasing threat, with both species considered to be of particular importance by the World Health Organization and the US Centers for Disease Control. The threat of antimicrobial resistance is exacerbated by the staggering difference in the speeds of development for the discovery and development of the antimicrobials versus resistance mechanisms. In the search for alternative strategies, modulation of host-pathogen interactions in general, and virulence inhibition in particular, have drawn substantial attention. Unfortunately, these approaches require a fairly comprehensive understanding of virulence determinants. This requirement is complicated by the fact that enterococcal infection models generally require vertebrates, making them slow, expensive, and ethically problematic, particularly when considering the thousands of animals that would be needed for the early stages of experimentation. To address this problem, we developed the first high-throughput C. elegans-E. faecium infection model involving host death. Importantly, this model recapitulates many key aspects of murine peritonitis models, including utilizing similar virulence determinants. Additionally, host death is independent of peroxide production, unlike other E. faecium-C. elegans virulence models, which allows the assessment of other virulence factors. Using this system, we analyzed a panel of lab strains with deletions of targeted virulence factors. Although removal of certain virulence factors (e.g., Δfms15) was sufficient to affect virulence, multiple deletions were generally required to affect pathogenesis, suggesting that host-pathogen interactions are multifactorial. These data were corroborated by genomic analysis of selected isolates with high and low levels of virulence. We anticipate that this platform will be useful for identifying new treatments for E. faecium infection.
Keywords: C. elegans; E. faecium; high-throughput screen; host-pathogen interaction; virulence.
Copyright © 2021 Revtovich, Tjahjono, Singh, Hanson, Murray and Kirienko.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous