

The role of Cytochrome b6f in the control of steady-state photosynthesis: a conceptual and quantitative model
- PMID: 33999328
- PMCID: PMC8292351
- DOI: 10.1007/s11120-021-00840-4
The role of Cytochrome b6f in the control of steady-state photosynthesis: a conceptual and quantitative model
Abstract
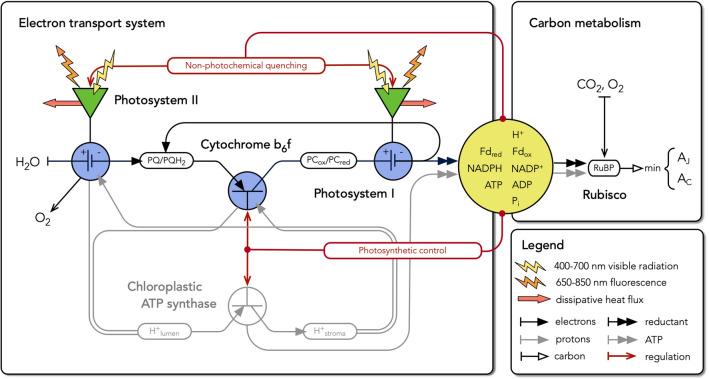
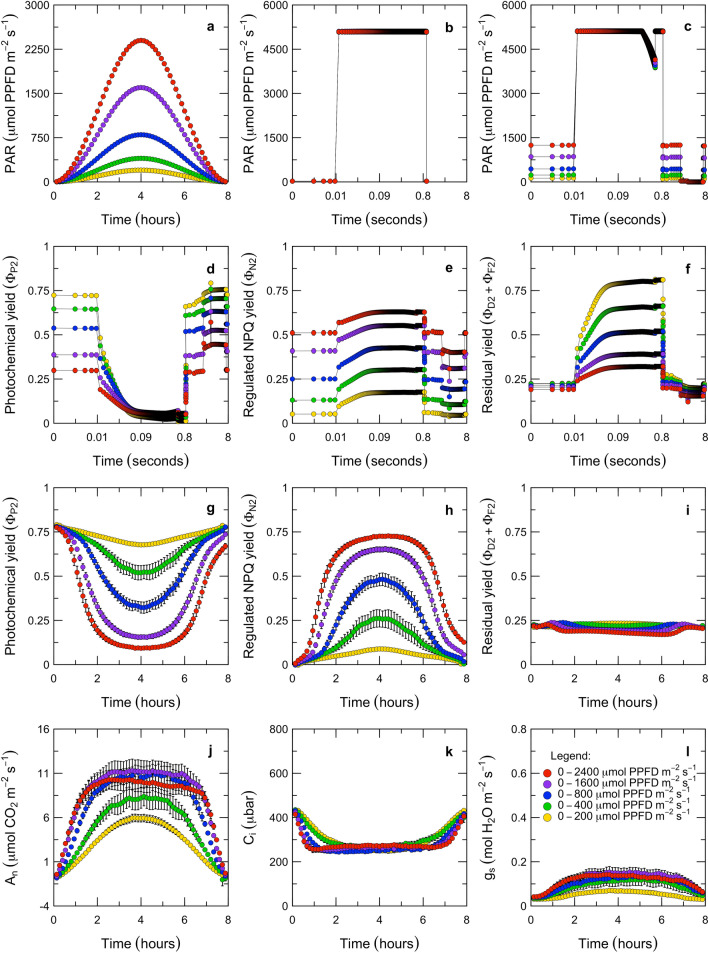
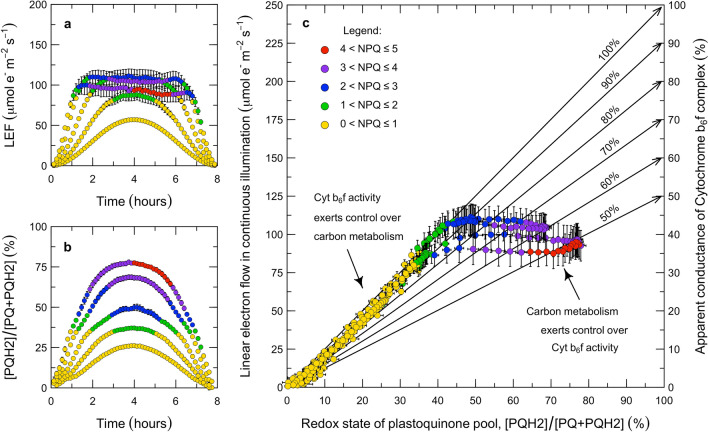
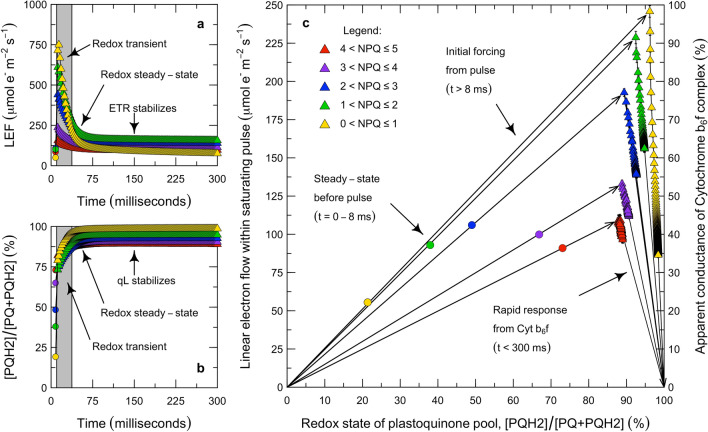
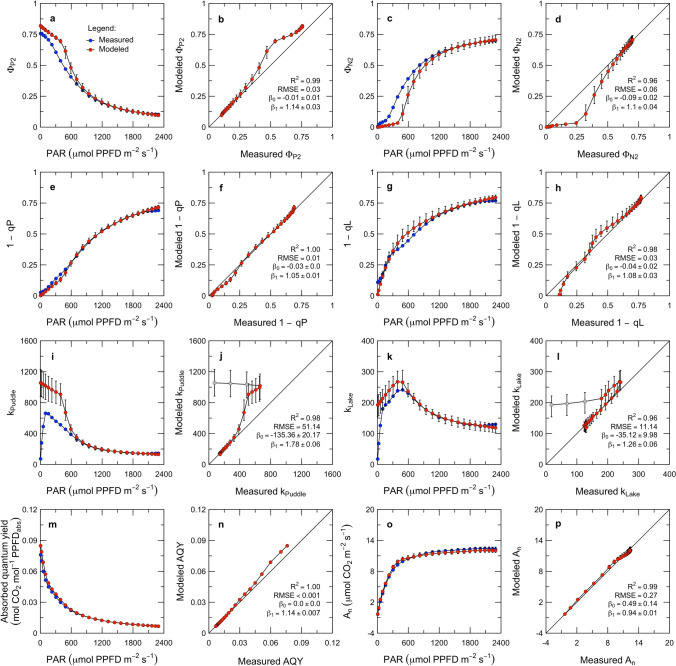
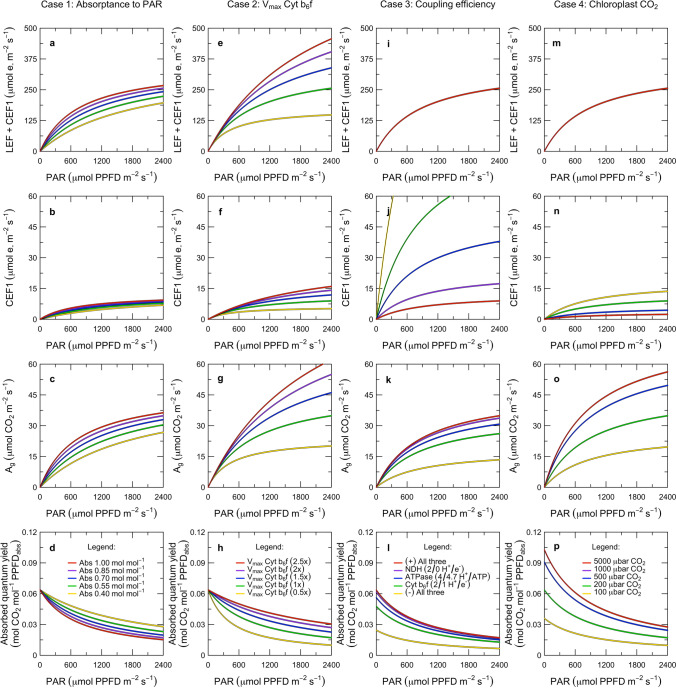
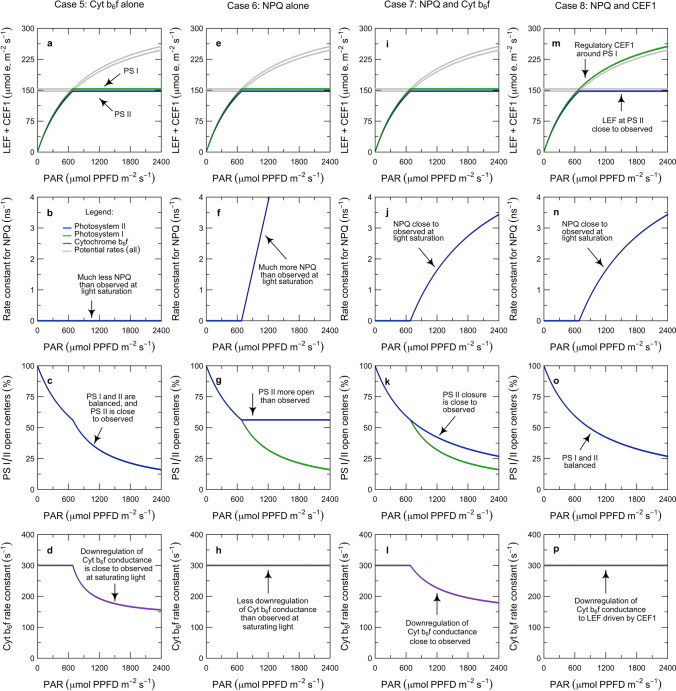
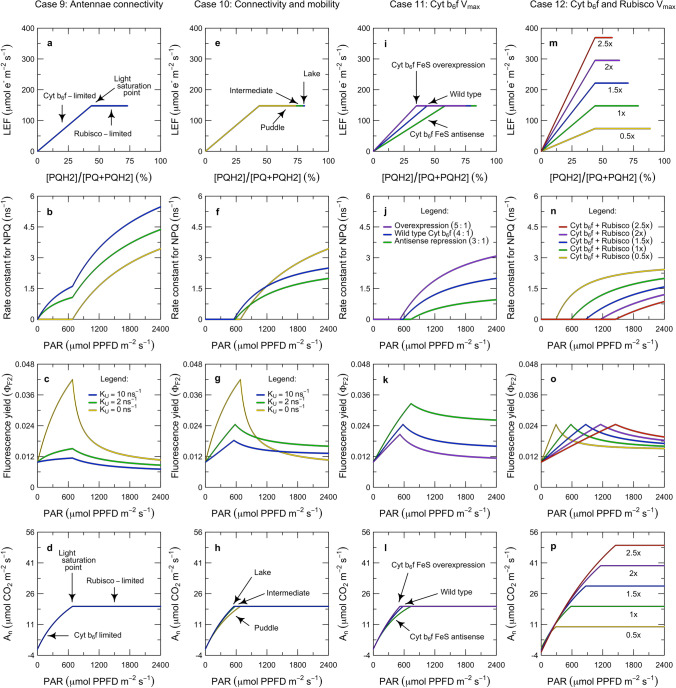
Here, we present a conceptual and quantitative model to describe the role of the Cytochrome [Formula: see text] complex in controlling steady-state electron transport in [Formula: see text] leaves. The model is based on new experimental methods to diagnose the maximum activity of Cyt [Formula: see text] in vivo, and to identify conditions under which photosynthetic control of Cyt [Formula: see text] is active or relaxed. With these approaches, we demonstrate that Cyt [Formula: see text] controls the trade-off between the speed and efficiency of electron transport under limiting light, and functions as a metabolic switch that transfers control to carbon metabolism under saturating light. We also present evidence that the onset of photosynthetic control of Cyt [Formula: see text] occurs within milliseconds of exposure to saturating light, much more quickly than the induction of non-photochemical quenching. We propose that photosynthetic control is the primary means of photoprotection and functions to manage excitation pressure, whereas non-photochemical quenching functions to manage excitation balance. We use these findings to extend the Farquhar et al. (Planta 149:78-90, 1980) model of [Formula: see text] photosynthesis to include a mechanistic description of the electron transport system. This framework relates the light captured by PS I and PS II to the energy and mass fluxes linking the photoacts with Cyt [Formula: see text], the ATP synthase, and Rubisco. It enables quantitative interpretation of pulse-amplitude modulated fluorometry and gas-exchange measurements, providing a new basis for analyzing how the electron transport system coordinates the supply of Fd, NADPH, and ATP with the dynamic demands of carbon metabolism, how efficient use of light is achieved under limiting light, and how photoprotection is achieved under saturating light. The model is designed to support forward as well as inverse applications. It can either be used in a stand-alone mode at the leaf-level or coupled to other models that resolve finer-scale or coarser-scale phenomena.
Keywords: Cytochrome; Electron transport; Model; Photosystem I; Photosystem II; Rubisco.
© 2021. The Author(s).
Conflict of interest statement
The authors have no relevant financial or non-financial interests to disclose.
Figures
References
-
- Anderson JM, Price GD, Chow WS, Hope AB, Badger MR. Reduced levels of cytochrome bf complex in transgenic tobacco leads to marked photochemical reduction of the plastoquinone pool, without significant change in acclimation to irradiance. Photosynth Res. 1997;53:215–227. doi: 10.1023/A:1005856615915. - DOI
-
- Avenson TJ, Saathoff AJ (2018) Sub-saturating multiphase flash irradiances to estimate maximum fluorescence yield. In: Covshoff S (ed) Photosynthesis: methods and protocols. Humana Press, New York, pp 105–120 - PubMed
-
- Baker NR, Oxborough K. Chlorophyll fluorescence as a probe of photosynthetic productivity. In: Papageorgiou GC, Govindjee, editors. Chlorophyll a fluorescence: a signature of photosynthesis. Berlin: Springer; 2004. pp. 65–82.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
