

DEFECTIVE EMBRYO AND MERISTEMS genes are required for cell division and gamete viability in Arabidopsis
- PMID: 33999950
- PMCID: PMC8158957
- DOI: 10.1371/journal.pgen.1009561
DEFECTIVE EMBRYO AND MERISTEMS genes are required for cell division and gamete viability in Arabidopsis
Abstract
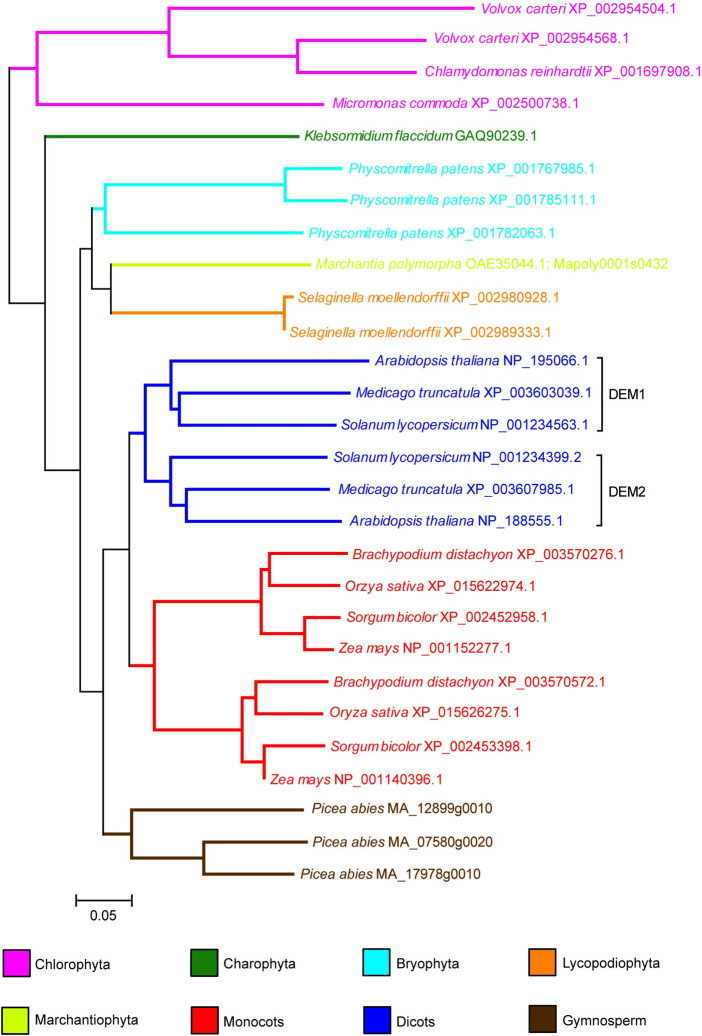
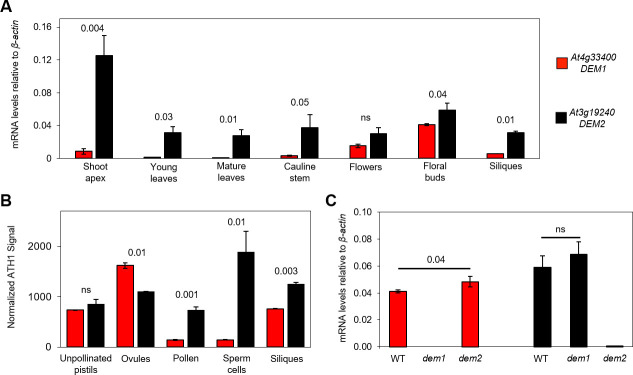
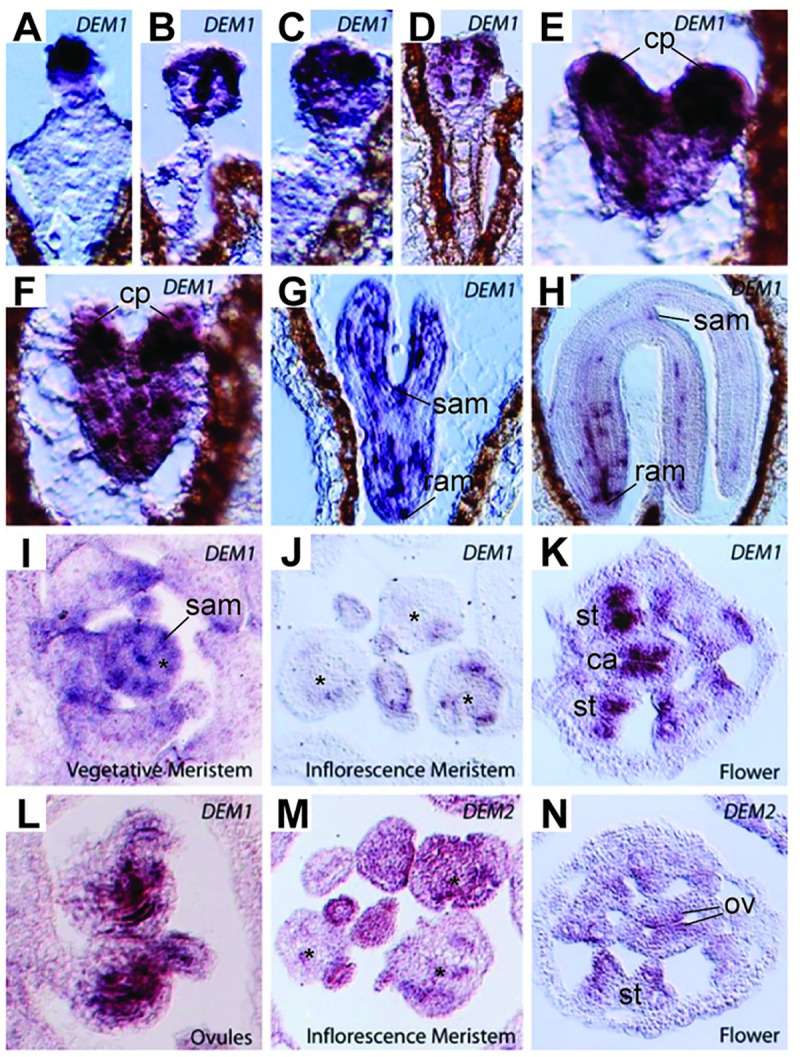
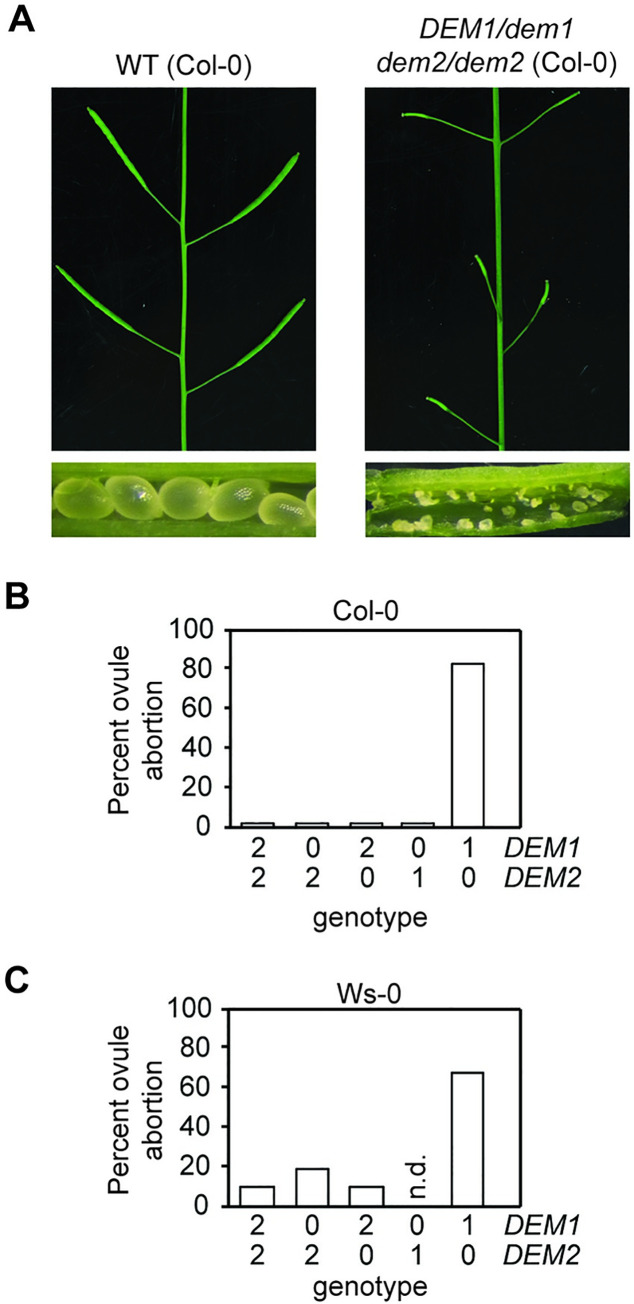
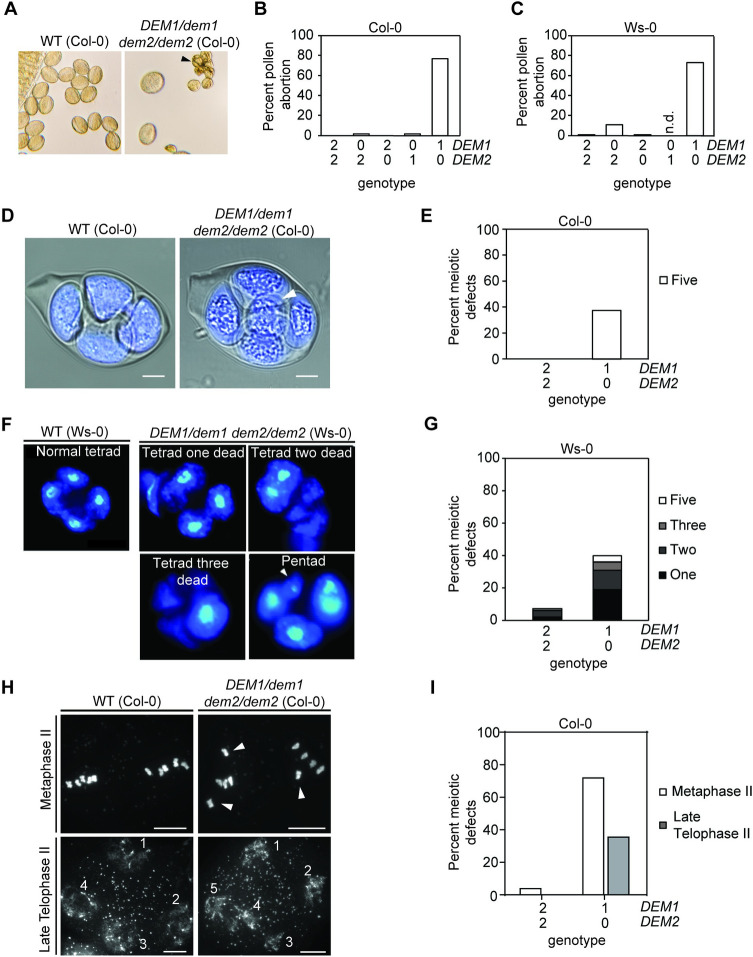
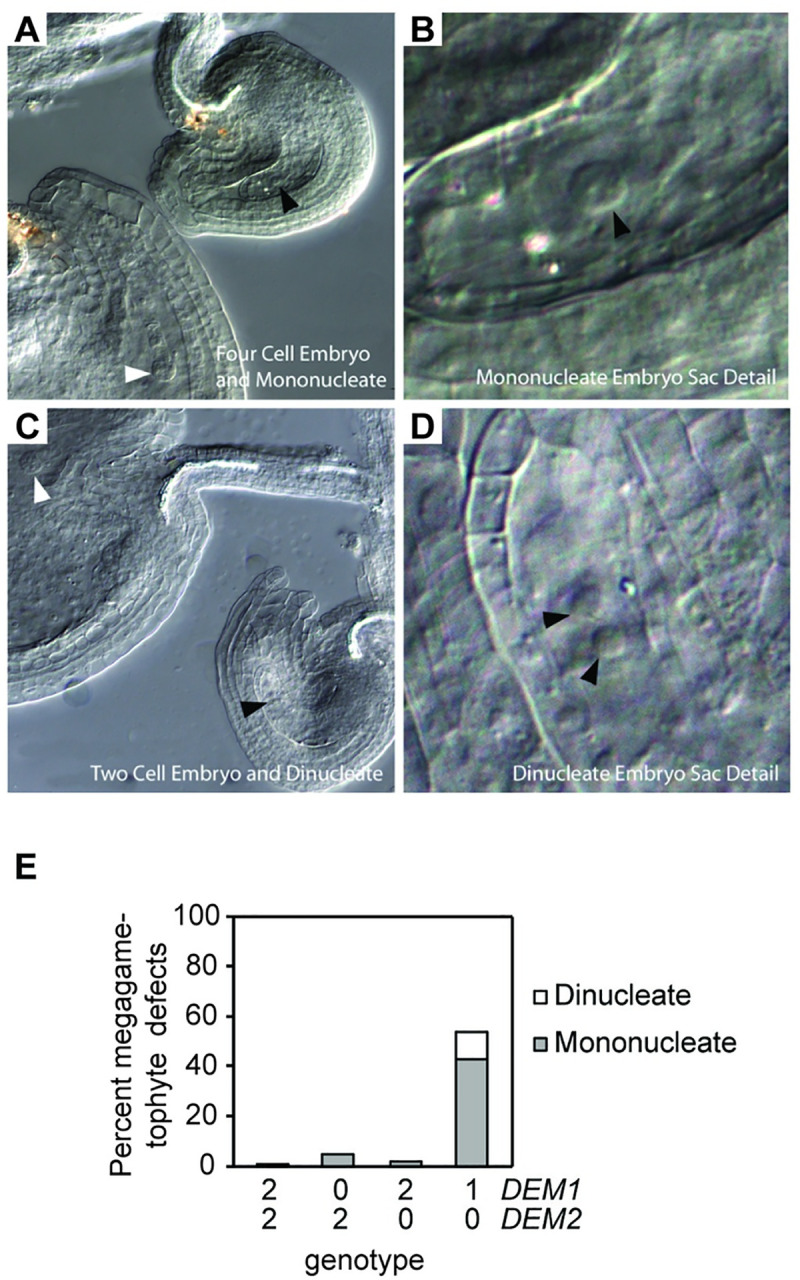
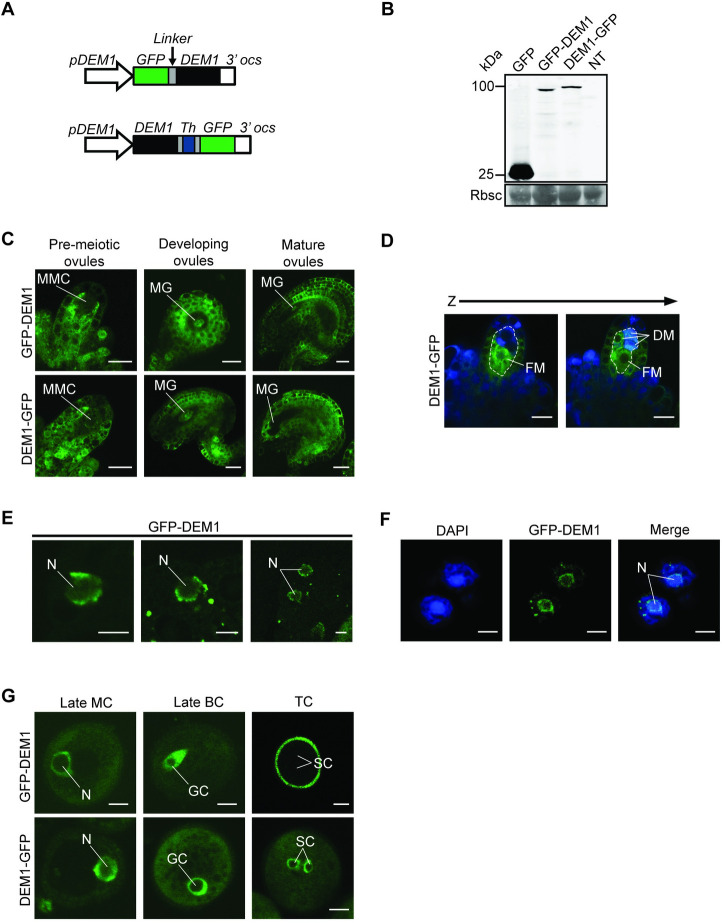
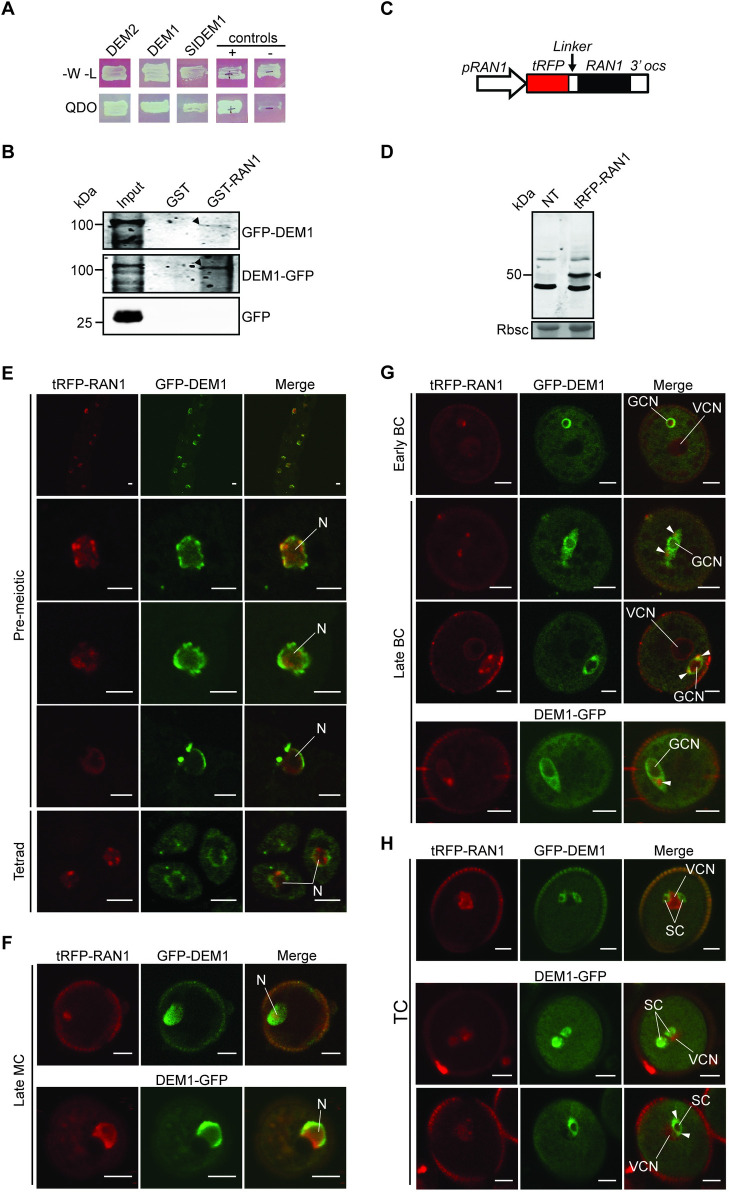
The DEFECTIVE EMBRYO AND MERISTEMS 1 (DEM1) gene encodes a protein of unknown biochemical function required for meristem formation and seedling development in tomato, but it was unclear whether DEM1's primary role was in cell division or alternatively, in defining the identity of meristematic cells. Genome sequence analysis indicates that flowering plants possess at least two DEM genes. Arabidopsis has two DEM genes, DEM1 and DEM2, which we show are expressed in developing embryos and meristems in a punctate pattern that is typical of genes involved in cell division. Homozygous dem1 dem2 double mutants were not recovered, and plants carrying a single functional DEM1 allele and no functional copies of DEM2, i.e. DEM1/dem1 dem2/dem2 plants, exhibit normal development through to the time of flowering but during male reproductive development, chromosomes fail to align on the metaphase plate at meiosis II and result in abnormal numbers of daughter cells following meiosis. Additionally, these plants show defects in both pollen and embryo sac development, and produce defective male and female gametes. In contrast, dem1/dem1 DEM2/dem2 plants showed normal levels of fertility, indicating that DEM2 plays a more important role than DEM1 in gamete viability. The increased importance of DEM2 in gamete viability correlated with higher mRNA levels of DEM2 compared to DEM1 in most tissues examined and particularly in the vegetative shoot apex, developing siliques, pollen and sperm. We also demonstrate that gamete viability depends not only on the number of functional DEM alleles inherited following meiosis, but also on the number of functional DEM alleles in the parent plant that undergoes meiosis. Furthermore, DEM1 interacts with RAS-RELATED NUCLEAR PROTEIN 1 (RAN1) in yeast two-hybrid and pull-down binding assays, and we show that fluorescent proteins fused to DEM1 and RAN1 co-localize transiently during male meiosis and pollen development. In eukaryotes, RAN is a highly conserved GTPase that plays key roles in cell cycle progression, spindle assembly during cell division, reformation of the nuclear envelope following cell division, and nucleocytoplasmic transport. Our results demonstrate that DEM proteins play an essential role in cell division in plants, most likely through an interaction with RAN1.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Qiu Y-L, Taylor AB, McManus HA. Evolution of the life cycle in land plants. Journal of Systematics and Evolution. 2012;50(3):171–94. 10.1111/j.1759-6831.2012.00188.x - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
