

Transcription Factor Dynamics
- PMID: 34001530
- PMCID: PMC8559544
- DOI: 10.1101/cshperspect.a040949
Transcription Factor Dynamics
Abstract
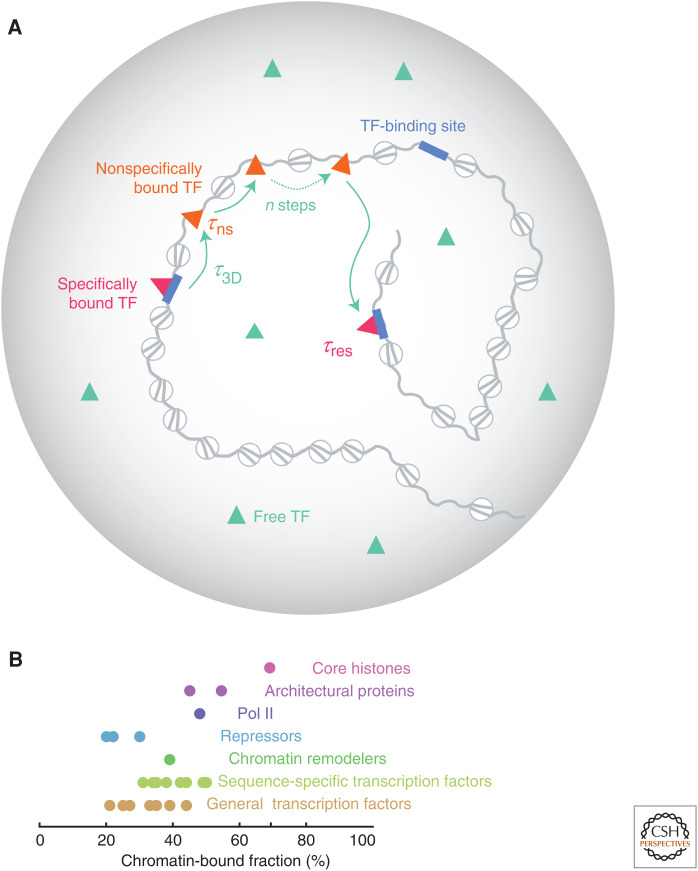
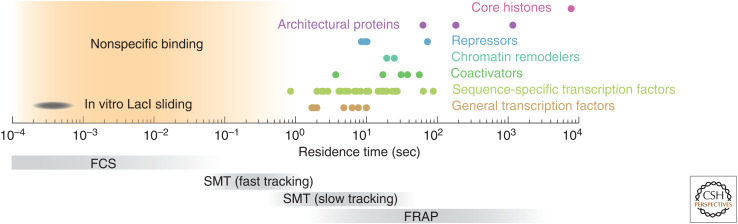
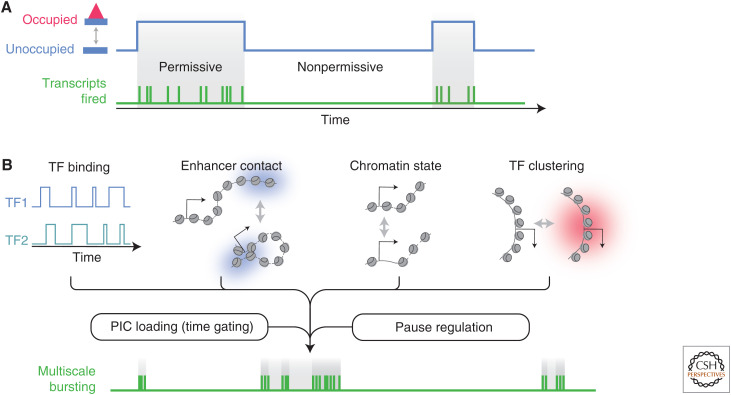
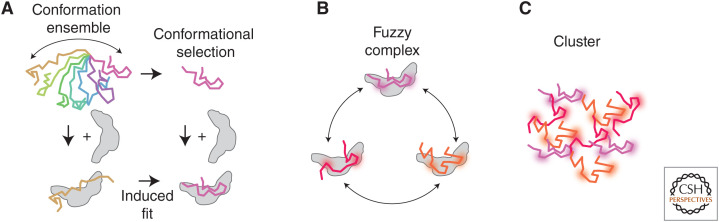
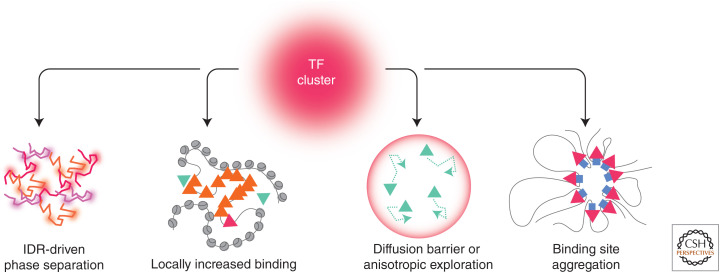
To predict transcription, one needs a mechanistic understanding of how the numerous required transcription factors (TFs) explore the nuclear space to find their target genes, assemble, cooperate, and compete with one another. Advances in fluorescence microscopy have made it possible to visualize real-time TF dynamics in living cells, leading to two intriguing observations: first, most TFs contact chromatin only transiently; and second, TFs can assemble into clusters through their intrinsically disordered regions. These findings suggest that highly dynamic events and spatially structured nuclear microenvironments might play key roles in transcription regulation that are not yet fully understood. The emerging model is that while some promoters directly convert TF-binding events into on/off cycles of transcription, many others apply complex regulatory layers that ultimately lead to diverse phenotypic outputs. Cracking this kinetic code is an ongoing and challenging task that is made possible by combining innovative imaging approaches with biophysical models.
Copyright © 2021 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous