

Genomic basis of parallel adaptation varies with divergence in Arabidopsis and its relatives
- PMID: 34001609
- PMCID: PMC8166048
- DOI: 10.1073/pnas.2022713118
Genomic basis of parallel adaptation varies with divergence in Arabidopsis and its relatives
Abstract
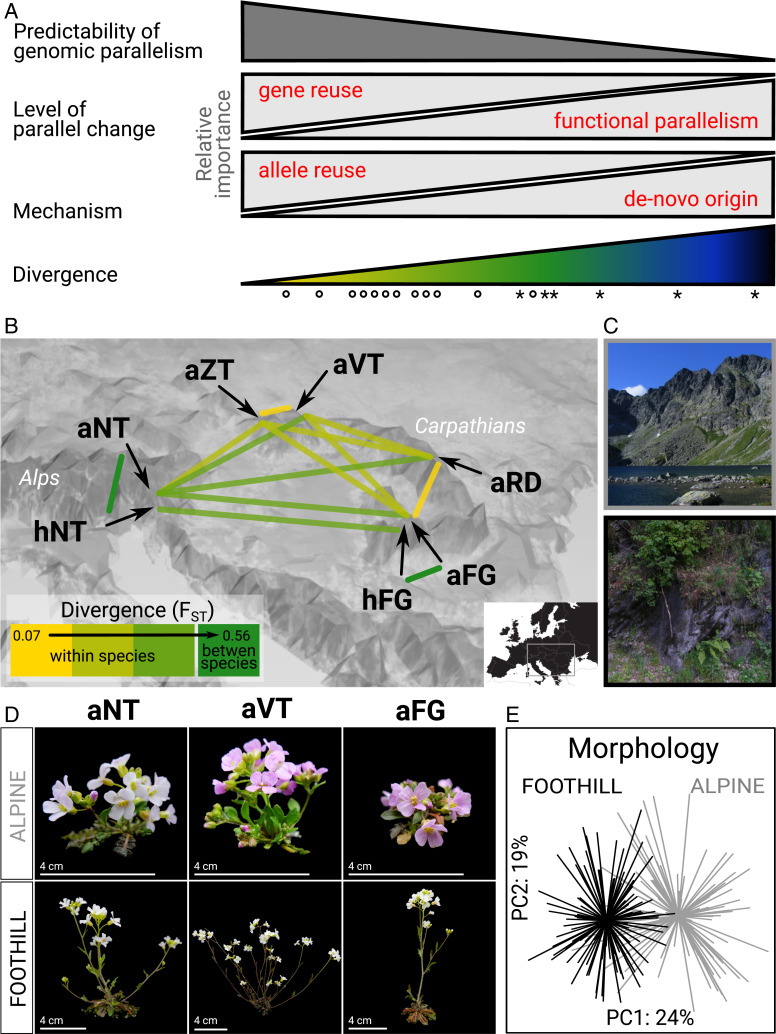
Parallel adaptation provides valuable insight into the predictability of evolutionary change through replicated natural experiments. A steadily increasing number of studies have demonstrated genomic parallelism, yet the magnitude of this parallelism varies depending on whether populations, species, or genera are compared. This led us to hypothesize that the magnitude of genomic parallelism scales with genetic divergence between lineages, but whether this is the case and the underlying evolutionary processes remain unknown. Here, we resequenced seven parallel lineages of two Arabidopsis species, which repeatedly adapted to challenging alpine environments. By combining genome-wide divergence scans with model-based approaches, we detected a suite of 151 genes that show parallel signatures of positive selection associated with alpine colonization, involved in response to cold, high radiation, short season, herbivores, and pathogens. We complemented these parallel candidates with published gene lists from five additional alpine Brassicaceae and tested our hypothesis on a broad scale spanning ∼0.02 to 18 My of divergence. Indeed, we found quantitatively variable genomic parallelism whose extent significantly decreased with increasing divergence between the compared lineages. We further modeled parallel evolution over the Arabidopsis candidate genes and showed that a decreasing probability of repeated selection on the same standing or introgressed alleles drives the observed pattern of divergence-dependent parallelism. We therefore conclude that genetic divergence between populations, species, and genera, affecting the pool of shared variants, is an important factor in the predictability of genome evolution.
Keywords: Arabidopsis; alpine adaptation; evolution; genomics; parallelism.
Conflict of interest statement
The authors declare no competing interest.
Figures



References
-
- Blount Z. D., Lenski R. E., Losos J. B., Contingency and determinism in evolution: Replaying life’s tape. Science 362, eaam5979 (2018). - PubMed
-
- Gould S. J., Wonderful Life : The Burgess Shale and the Nature of History (Norton, 1989).
-
- Agrawal A. A., Toward a predictive framework for convergent evolution: Integrating natural history, genetic mechanisms, and consequences for the diversity of Life. Am. Nat. 190 (S1), S1–S12 (2017). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
