

PRMT6 activates cyclin D1 expression in conjunction with the transcription factor LEF1
- PMID: 34001852
- PMCID: PMC8129428
- DOI: 10.1038/s41389-021-00332-z
PRMT6 activates cyclin D1 expression in conjunction with the transcription factor LEF1
Abstract
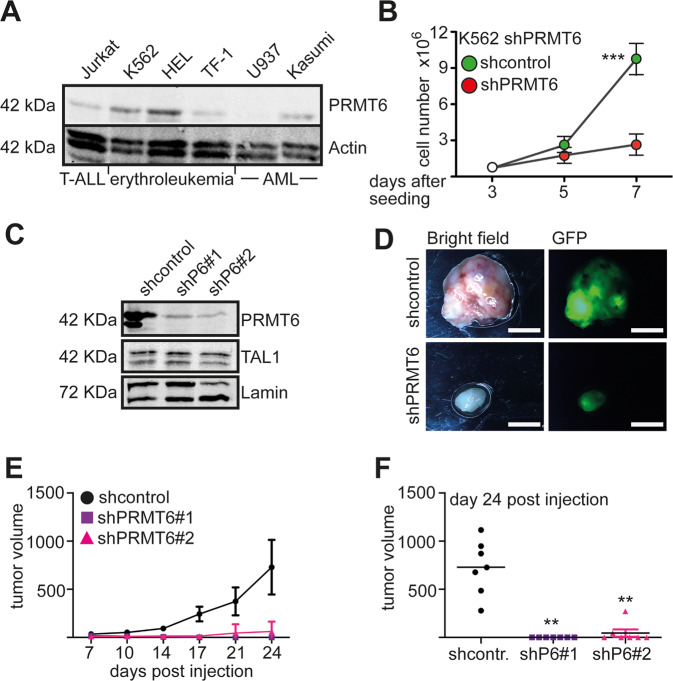
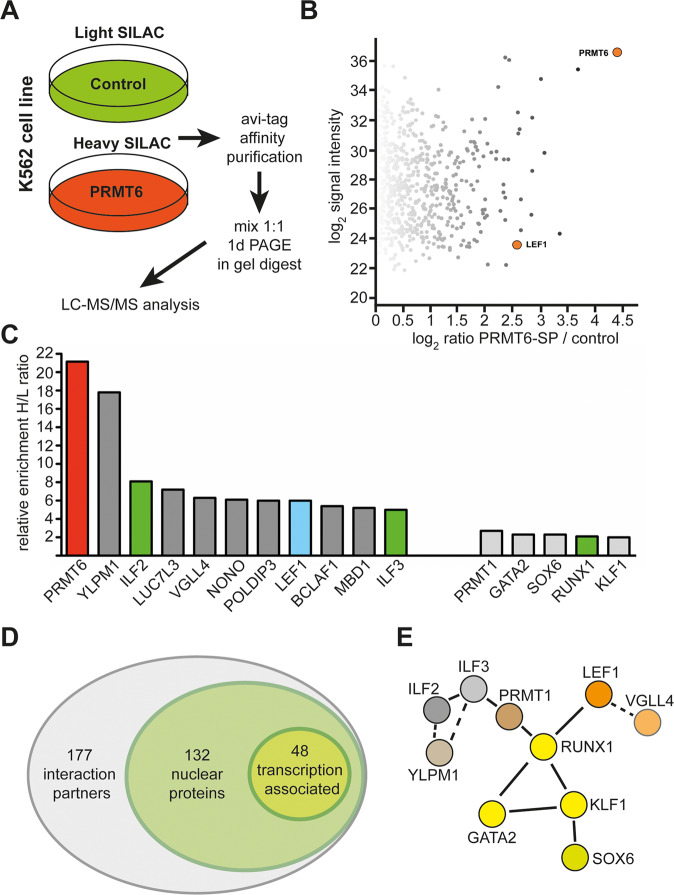
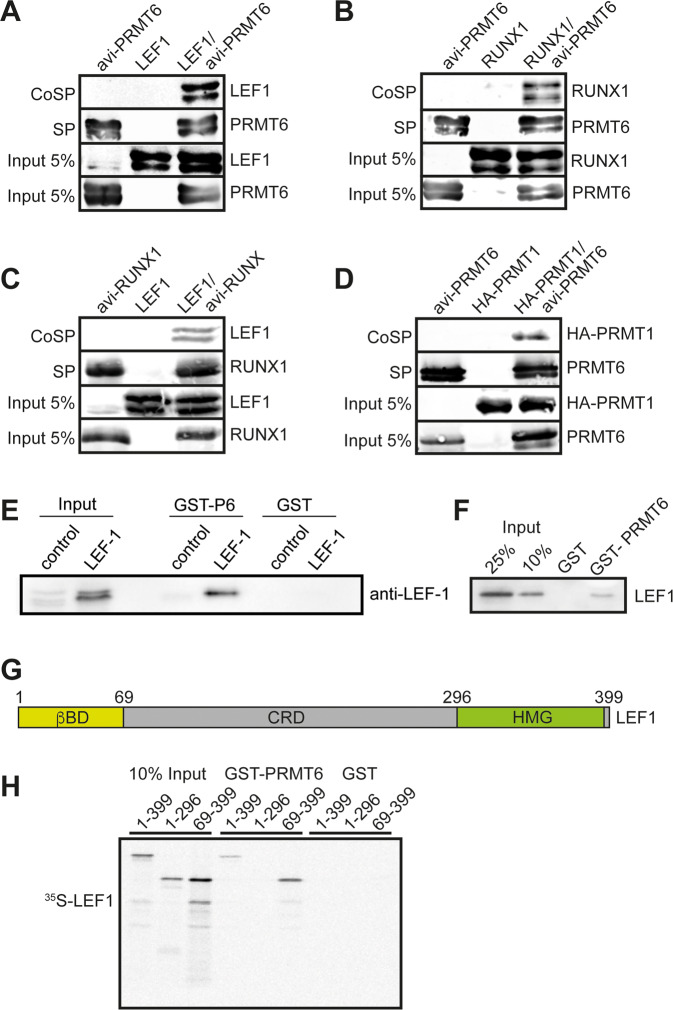
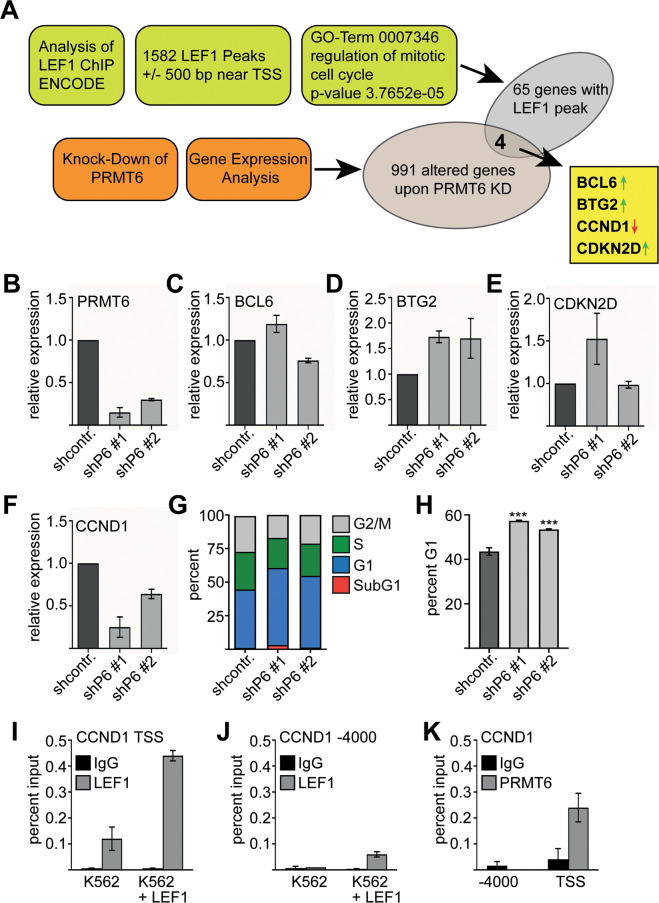
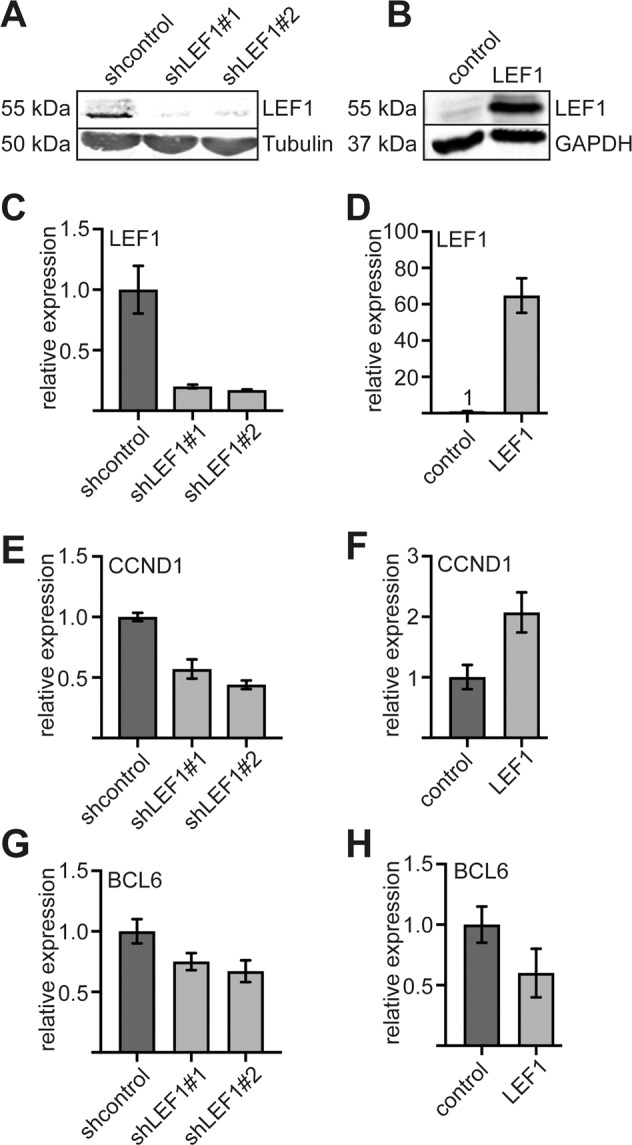
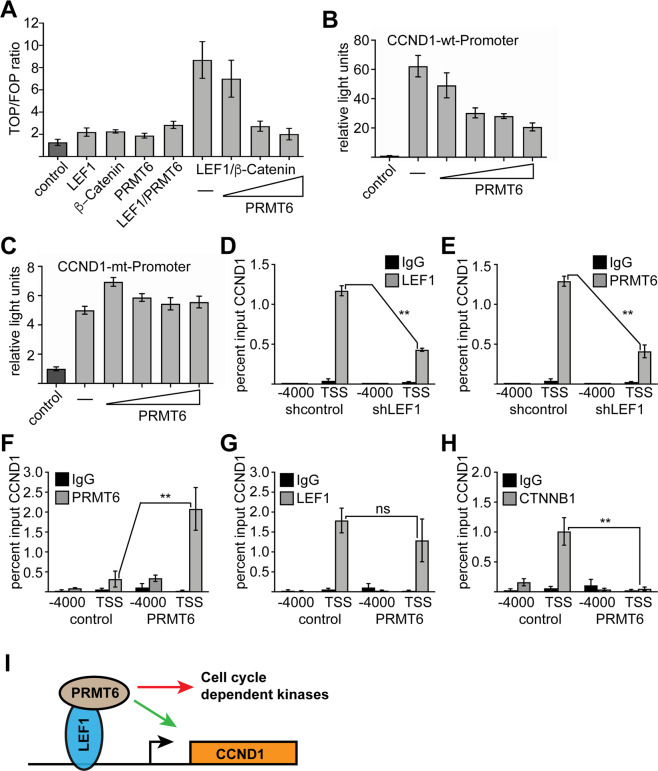
The establishment of cell type specific gene expression by transcription factors and their epigenetic cofactors is central for cell fate decisions. Protein arginine methyltransferase 6 (PRMT6) is an epigenetic regulator of gene expression mainly through methylating arginines at histone H3. This way it influences cellular differentiation and proliferation. PRMT6 lacks DNA-binding capability but is recruited by transcription factors to regulate gene expression. However, currently only a limited number of transcription factors have been identified, which facilitate recruitment of PRMT6 to key cell cycle related target genes. Here, we show that LEF1 contributes to the recruitment of PRMT6 to the central cell cycle regulator CCND1 (Cyclin D1). We identified LEF1 as an interaction partner of PRMT6. Knockdown of LEF1 or PRMT6 reduces CCND1 expression. This is in line with our observation that knockdown of PRMT6 increases the number of cells in G1 phase of the cell cycle and decreases proliferation. These results improve the understanding of PRMT6 activity in cell cycle regulation. We expect that these insights will foster the rational development and usage of specific PRMT6 inhibitors for cancer therapy.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Contributions of the histone arginine methyltransferase PRMT6 to the epigenetic function of RUNX1.Crit Rev Eukaryot Gene Expr. 2013;23(3):265-74. doi: 10.1615/critreveukaryotgeneexpr.2013007527. Crit Rev Eukaryot Gene Expr. 2013. PMID: 23879542 Review.
-
Protein arginine methyltransferase 6 controls erythroid gene expression and differentiation of human CD34+ progenitor cells.Haematologica. 2018 Jan;103(1):18-29. doi: 10.3324/haematol.2017.174516. Epub 2017 Oct 12. Haematologica. 2018. PMID: 29025910 Free PMC article.
-
Protein arginine methyltransferase 6 mediates cardiac hypertrophy by differential regulation of histone H3 arginine methylation.Heliyon. 2020 May 12;6(5):e03864. doi: 10.1016/j.heliyon.2020.e03864. eCollection 2020 May. Heliyon. 2020. PMID: 32420474 Free PMC article.
-
Assessment of PRMT6-dependent alternative splicing in pluripotent and differentiating NT2/D1 cells.Life Sci Alliance. 2025 Feb 3;8(4):e202402946. doi: 10.26508/lsa.202402946. Print 2025 Apr. Life Sci Alliance. 2025. PMID: 39900436 Free PMC article.
-
The Emerging Role of PRMT6 in Cancer.Front Oncol. 2022 Mar 4;12:841381. doi: 10.3389/fonc.2022.841381. eCollection 2022. Front Oncol. 2022. PMID: 35311114 Free PMC article. Review.
Cited by
-
PRMT6-mediated transcriptional activation of ythdf2 promotes glioblastoma migration, invasion, and emt via the wnt-β-catenin pathway.J Exp Clin Cancer Res. 2024 Apr 18;43(1):116. doi: 10.1186/s13046-024-03038-3. J Exp Clin Cancer Res. 2024. PMID: 38637831 Free PMC article.
-
Wnt/β-Catenin-Signaling Modulates Megakaryopoiesis at the Megakaryocyte-Erythrocyte Progenitor Stage in the Hematopoietic System.Cells. 2023 Dec 4;12(23):2765. doi: 10.3390/cells12232765. Cells. 2023. PMID: 38067194 Free PMC article.
-
PRMT6-CDC20 facilitates glioblastoma progression via the degradation of CDKN1B.Oncogene. 2023 Mar;42(14):1088-1100. doi: 10.1038/s41388-023-02624-7. Epub 2023 Feb 15. Oncogene. 2023. PMID: 36792756 Free PMC article.
-
Carcinogenesis promotion in oral squamous cell carcinoma: KDM4A complex-mediated gene transcriptional suppression by LEF1.Cell Death Dis. 2023 Aug 8;14(8):510. doi: 10.1038/s41419-023-06024-3. Cell Death Dis. 2023. PMID: 37553362 Free PMC article.
-
Hetero-oligomeric interaction as a new regulatory mechanism for protein arginine methyltransferases.Biochem Soc Trans. 2024 Oct 30;52(5):2193-2201. doi: 10.1042/BST20240242. Biochem Soc Trans. 2024. PMID: 39324605 Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
