

Proline codon pair selection determines ribosome pausing strength and translation efficiency in bacteria
- PMID: 34002016
- PMCID: PMC8129111
- DOI: 10.1038/s42003-021-02115-z
Proline codon pair selection determines ribosome pausing strength and translation efficiency in bacteria
Abstract
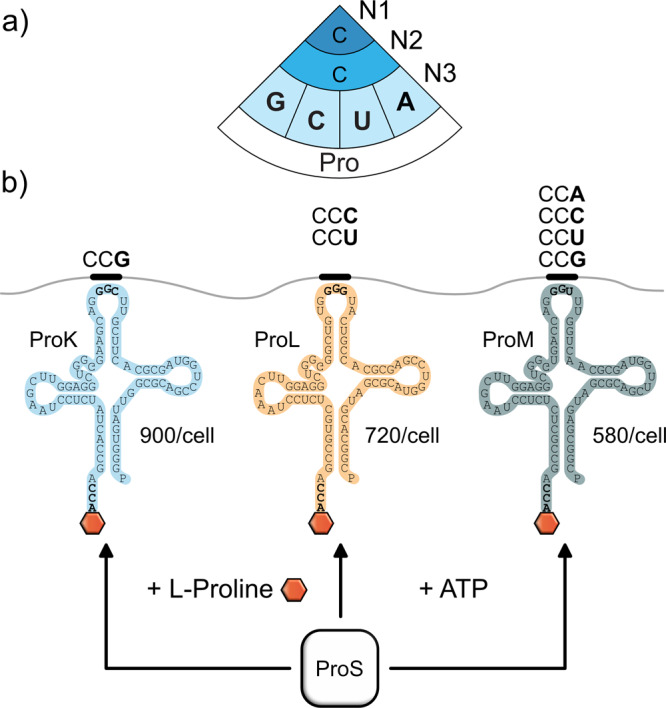
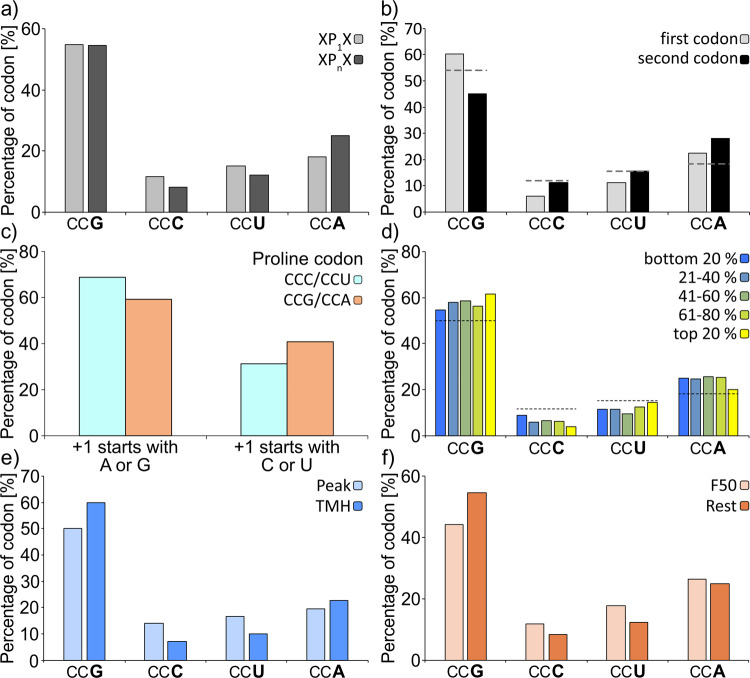
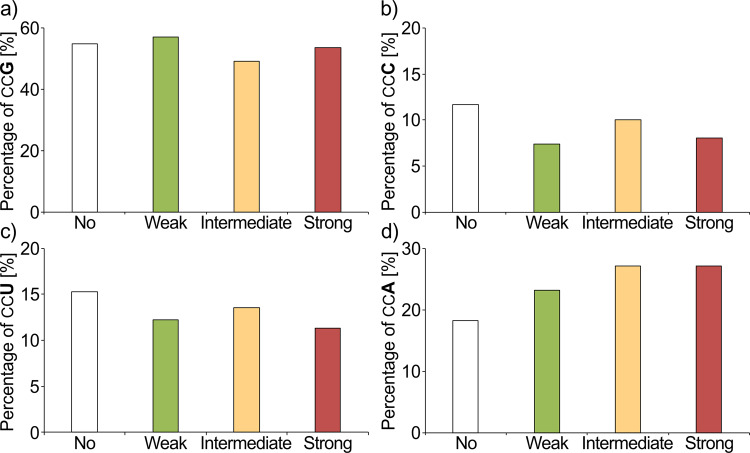
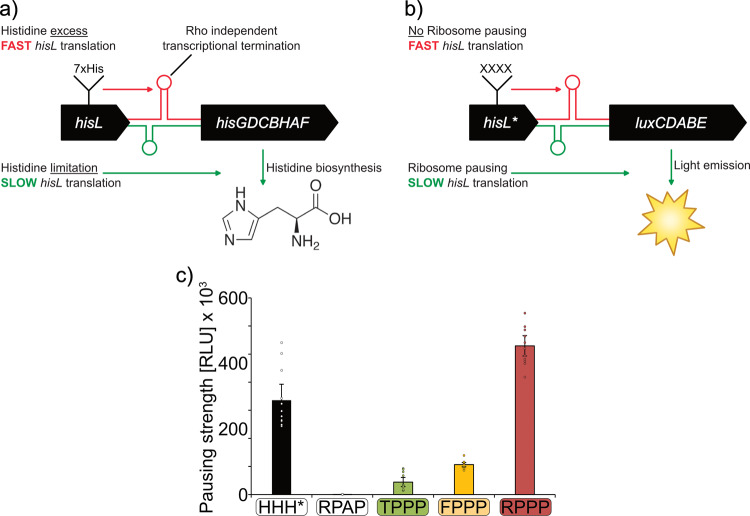
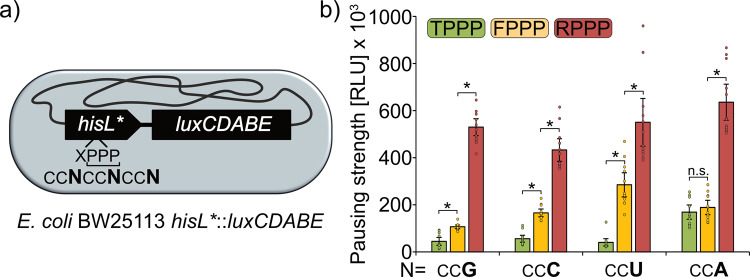
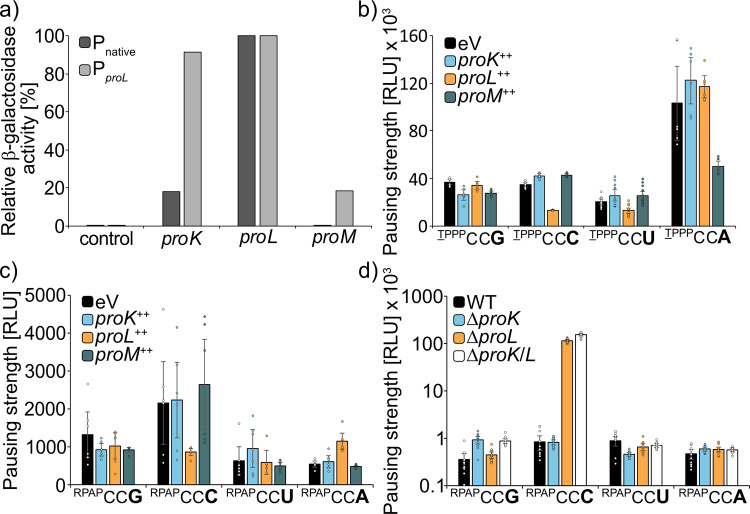
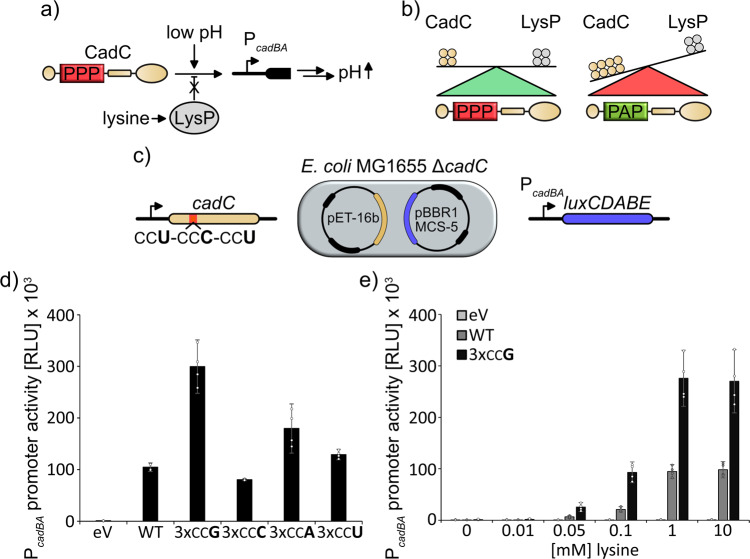
The speed of mRNA translation depends in part on the amino acid to be incorporated into the nascent chain. Peptide bond formation is especially slow with proline and two adjacent prolines can even cause ribosome stalling. While previous studies focused on how the amino acid context of a Pro-Pro motif determines the stalling strength, we extend this question to the mRNA level. Bioinformatics analysis of the Escherichia coli genome revealed significantly differing codon usage between single and consecutive prolines. We therefore developed a luminescence reporter to detect ribosome pausing in living cells, enabling us to dissect the roles of codon choice and tRNA selection as well as to explain the genome scale observations. Specifically, we found a strong selective pressure against CCC/U-C, a sequon causing ribosomal frameshifting even under wild-type conditions. On the other hand, translation efficiency as positive evolutionary driving force led to an overrepresentation of CCG. This codon is not only translated the fastest, but the corresponding prolyl-tRNA reaches almost saturating levels. By contrast, CCA, for which the cognate prolyl-tRNA amounts are limiting, is used to regulate pausing strength. Thus, codon selection both in discrete positions but especially in proline codon pairs can tune protein copy numbers.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Spatial Distribution and Ribosome-Binding Dynamics of EF-P in Live Escherichia coli.mBio. 2017 Jun 6;8(3):e00300-17. doi: 10.1128/mBio.00300-17. mBio. 2017. PMID: 28588135 Free PMC article.
-
Versatile Dual Reporter to Identify Ribosome Pausing Motifs Alleviated by Translation Elongation Factor P.ACS Synth Biol. 2024 Nov 15;13(11):3698-3710. doi: 10.1021/acssynbio.4c00534. Epub 2024 Oct 19. ACS Synth Biol. 2024. PMID: 39425678
-
Ribosome kinetics and aa-tRNA competition determine rate and fidelity of peptide synthesis.Comput Biol Chem. 2007 Oct;31(5-6):335-46. doi: 10.1016/j.compbiolchem.2007.07.003. Epub 2007 Aug 15. Comput Biol Chem. 2007. PMID: 17897886 Free PMC article.
-
Optimizing scaleup yield for protein production: Computationally Optimized DNA Assembly (CODA) and Translation Engineering.Biotechnol Annu Rev. 2007;13:27-42. doi: 10.1016/S1387-2656(07)13002-7. Biotechnol Annu Rev. 2007. PMID: 17875472 Review.
-
Elongation Factor P and the Control of Translation Elongation.Annu Rev Microbiol. 2017 Sep 8;71:117-131. doi: 10.1146/annurev-micro-090816-093629. Epub 2017 May 22. Annu Rev Microbiol. 2017. PMID: 28886684 Review.
Cited by
-
Evaluating Performance of Different RNA Secondary Structure Prediction Programs Using Self-cleaving Ribozymes.Genomics Proteomics Bioinformatics. 2024 Sep 13;22(3):qzae043. doi: 10.1093/gpbjnl/qzae043. Genomics Proteomics Bioinformatics. 2024. PMID: 39317944 Free PMC article.
-
Kinetic mechanism and determinants of EF-P recruitment to translating ribosomes.Nucleic Acids Res. 2024 Oct 28;52(19):11870-11883. doi: 10.1093/nar/gkae815. Nucleic Acids Res. 2024. PMID: 39315709 Free PMC article.
-
Synonymous and non-synonymous codon substitutions can alleviate dependence on GroEL for folding.Protein Sci. 2024 Aug;33(8):e5087. doi: 10.1002/pro.5087. Protein Sci. 2024. PMID: 39074255 Free PMC article.
-
Analysis of 3.5 million SARS-CoV-2 sequences reveals unique mutational trends with consistent nucleotide and codon frequencies.Virol J. 2023 Feb 17;20(1):31. doi: 10.1186/s12985-023-01982-8. Virol J. 2023. PMID: 36812119 Free PMC article.
-
VeloPro: A pipeline integrating Ribo-seq and AlphaFold deciphers association patterns between translation velocity and protein structure features.Imeta. 2023 Nov 19;2(4):e148. doi: 10.1002/imt2.148. eCollection 2023 Nov. Imeta. 2023. PMID: 38868219 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
