

RAD54 is essential for RAD51-mediated repair of meiotic DSB in Arabidopsis
- PMID: 34003859
- PMCID: PMC8162660
- DOI: 10.1371/journal.pgen.1008919
RAD54 is essential for RAD51-mediated repair of meiotic DSB in Arabidopsis
Abstract
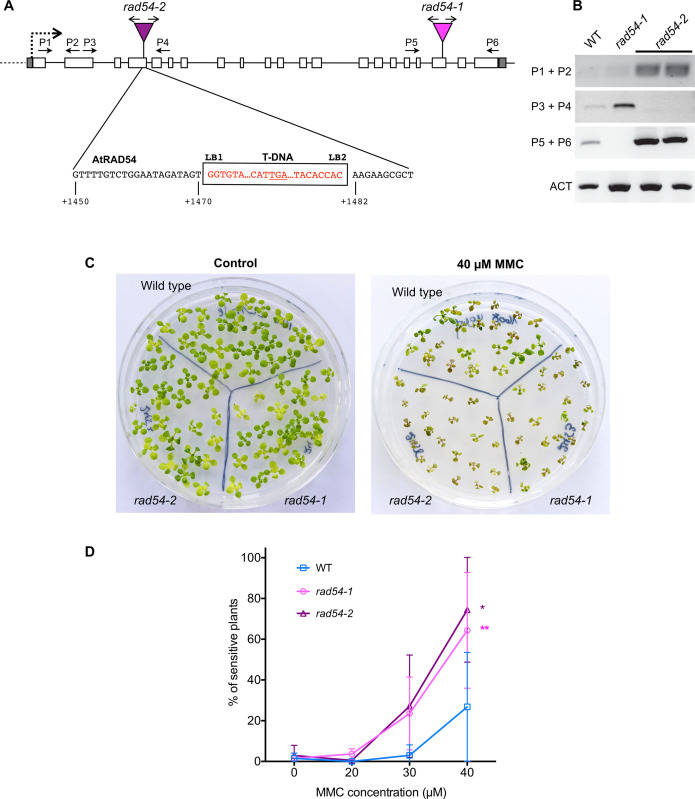
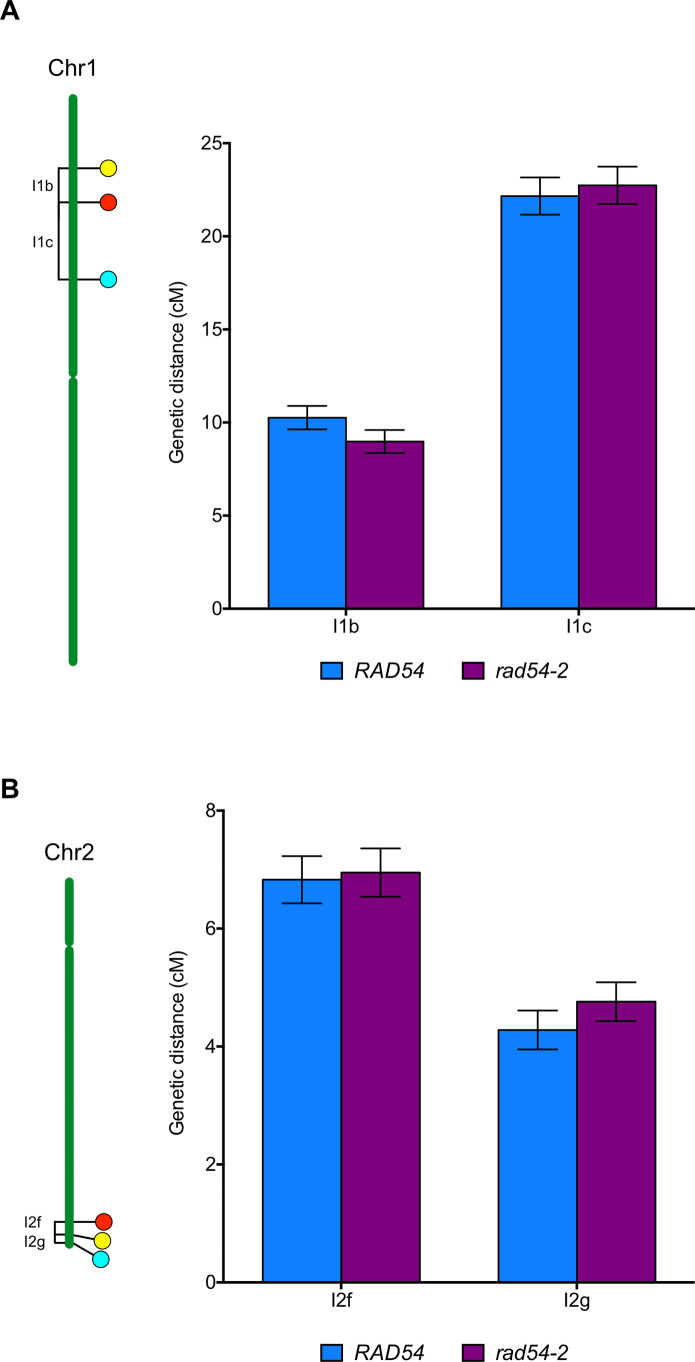
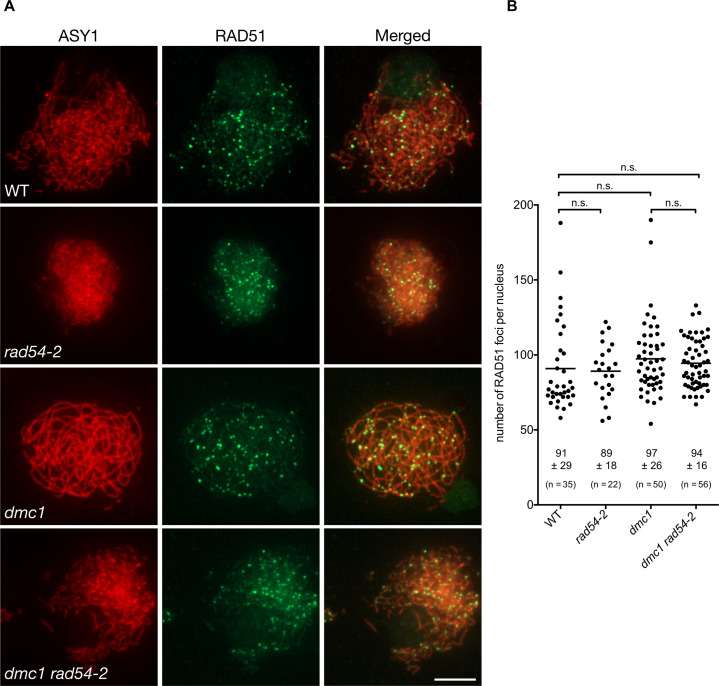
An essential component of the homologous recombination machinery in eukaryotes, the RAD54 protein is a member of the SWI2/SNF2 family of helicases with dsDNA-dependent ATPase, DNA translocase, DNA supercoiling and chromatin remodelling activities. It is a motor protein that translocates along dsDNA and performs multiple functions in homologous recombination. In particular, RAD54 is an essential cofactor for regulating RAD51 activity. It stabilizes the RAD51 nucleofilament, remodels nucleosomes, and stimulates the homology search and strand invasion activities of RAD51. Accordingly, deletion of RAD54 has dramatic consequences on DNA damage repair in mitotic cells. In contrast, its role in meiotic recombination is less clear. RAD54 is essential for meiotic recombination in Drosophila and C. elegans, but plays minor roles in yeast and mammals. We present here characterization of the roles of RAD54 in meiotic recombination in the model plant Arabidopsis thaliana. Absence of RAD54 has no detectable effect on meiotic recombination in otherwise wild-type plants but RAD54 becomes essential for meiotic DSB repair in absence of DMC1. In Arabidopsis, dmc1 mutants have an achiasmate meiosis, in which RAD51 repairs meiotic DSBs. Lack of RAD54 leads to meiotic chromosomal fragmentation in absence of DMC1. The action of RAD54 in meiotic RAD51 activity is thus mainly downstream of the role of RAD51 in supporting the activity of DMC1. Equivalent analyses show no effect on meiosis of combining dmc1 with the mutants of the RAD51-mediators RAD51B, RAD51D and XRCC2. RAD54 is thus required for repair of meiotic DSBs by RAD51 and the absence of meiotic phenotype in rad54 plants is a consequence of RAD51 playing a RAD54-independent supporting role to DMC1 in meiotic recombination.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



Similar articles
-
Repair of DNA double-strand breaks in plant meiosis: role of eukaryotic RecA recombinases and their modulators.Plant Reprod. 2023 Mar;36(1):17-41. doi: 10.1007/s00497-022-00443-6. Epub 2022 Jun 1. Plant Reprod. 2023. PMID: 35641832 Review.
-
MCM8 is required for a pathway of meiotic double-strand break repair independent of DMC1 in Arabidopsis thaliana.PLoS Genet. 2013;9(1):e1003165. doi: 10.1371/journal.pgen.1003165. Epub 2013 Jan 3. PLoS Genet. 2013. PMID: 23300481 Free PMC article.
-
The recombinases DMC1 and RAD51 are functionally and spatially separated during meiosis in Arabidopsis.Plant Cell. 2012 May;24(5):2058-70. doi: 10.1105/tpc.112.098459. Epub 2012 May 15. Plant Cell. 2012. PMID: 22589466 Free PMC article.
-
Meiotic recombination in Arabidopsis is catalysed by DMC1, with RAD51 playing a supporting role.PLoS Genet. 2013;9(9):e1003787. doi: 10.1371/journal.pgen.1003787. Epub 2013 Sep 26. PLoS Genet. 2013. PMID: 24086145 Free PMC article.
-
Roles of RecA homologues Rad51 and Dmc1 during meiotic recombination.Cytogenet Genome Res. 2004;107(3-4):201-7. doi: 10.1159/000080598. Cytogenet Genome Res. 2004. PMID: 15467365 Review.
Cited by
-
DMC1 attenuates RAD51-mediated recombination in Arabidopsis.PLoS Genet. 2022 Aug 25;18(8):e1010322. doi: 10.1371/journal.pgen.1010322. eCollection 2022 Aug. PLoS Genet. 2022. PMID: 36007010 Free PMC article.
-
Repair of DNA double-strand breaks in plant meiosis: role of eukaryotic RecA recombinases and their modulators.Plant Reprod. 2023 Mar;36(1):17-41. doi: 10.1007/s00497-022-00443-6. Epub 2022 Jun 1. Plant Reprod. 2023. PMID: 35641832 Review.
-
Homologous Recombination and Repair Functions Required for Mutagenicity during Yeast Meiosis.Genes (Basel). 2023 Oct 28;14(11):2017. doi: 10.3390/genes14112017. Genes (Basel). 2023. PMID: 38002960 Free PMC article.
-
FIGL1 attenuates meiotic interhomolog repair and is counteracted by the RAD51 paralog XRCC2 and the chromosome axis protein ASY1 during meiosis.New Phytol. 2024 Dec;244(6):2442-2457. doi: 10.1111/nph.20181. Epub 2024 Oct 17. New Phytol. 2024. PMID: 39420761 Free PMC article.
-
PlantCHRs: A comprehensive database of plant chromatin remodeling factors.Comput Struct Biotechnol J. 2023 Oct 5;21:4974-4987. doi: 10.1016/j.csbj.2023.10.005. eCollection 2023. Comput Struct Biotechnol J. 2023. PMID: 37867975 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
