

Small RNAs are modified with N-glycans and displayed on the surface of living cells
- PMID: 34004145
- PMCID: PMC9097497
- DOI: 10.1016/j.cell.2021.04.023
Small RNAs are modified with N-glycans and displayed on the surface of living cells
Erratum in
-
Small RNAs are modified with N-glycans and displayed on the surface of living cells.Cell. 2025 Aug 7;188(16):4470. doi: 10.1016/j.cell.2025.07.019. Epub 2025 Jul 22. Cell. 2025. PMID: 40695275 Free PMC article. No abstract available.
Abstract
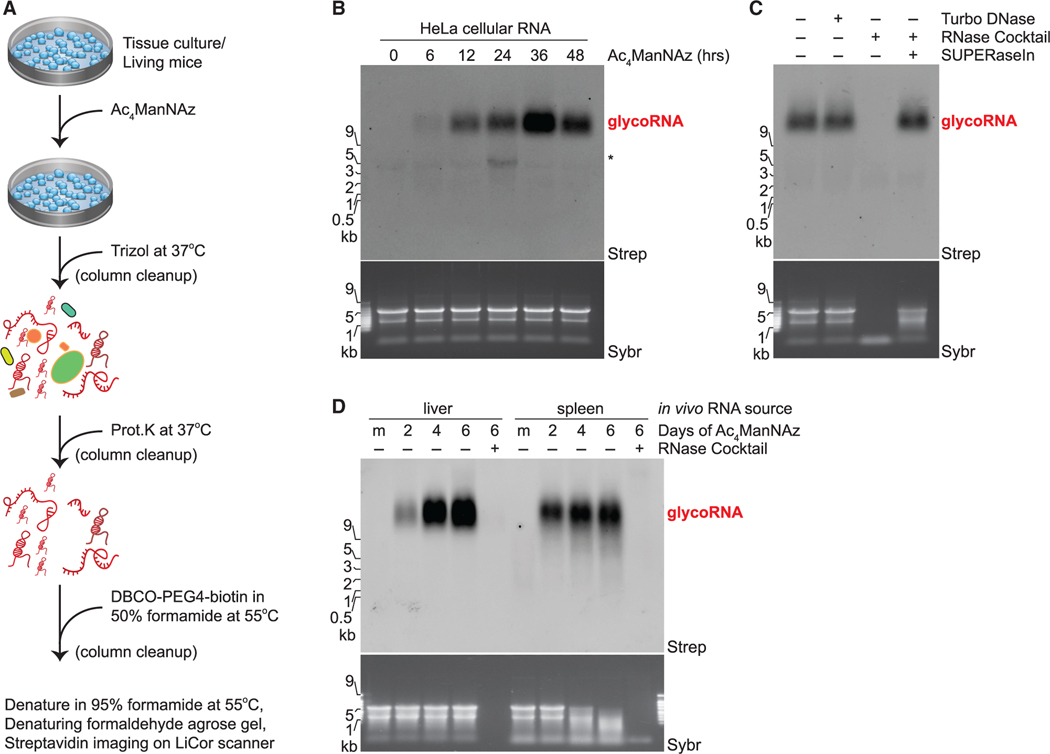
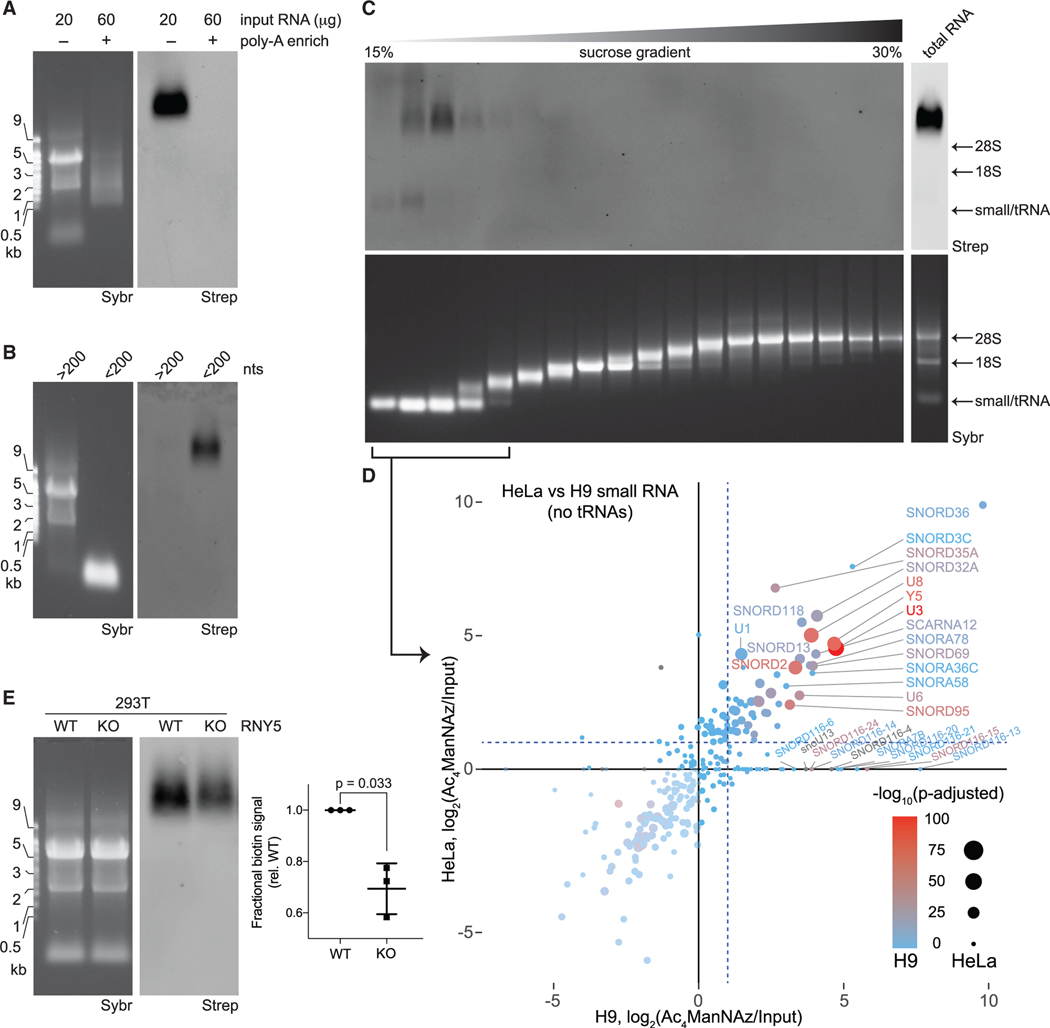
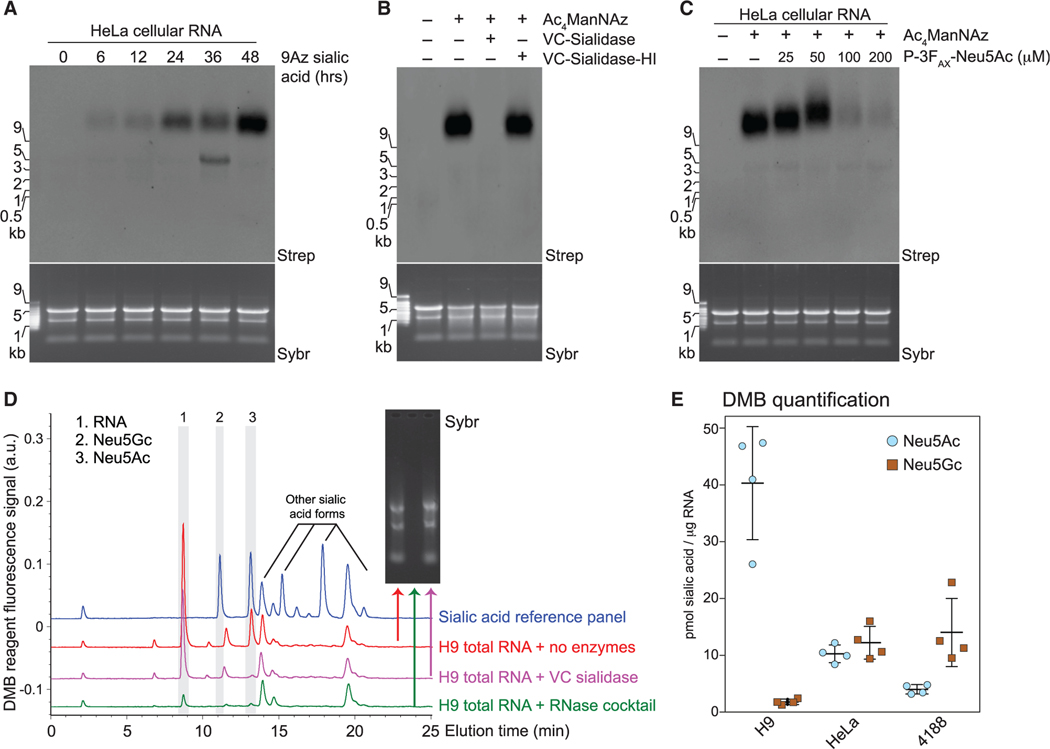
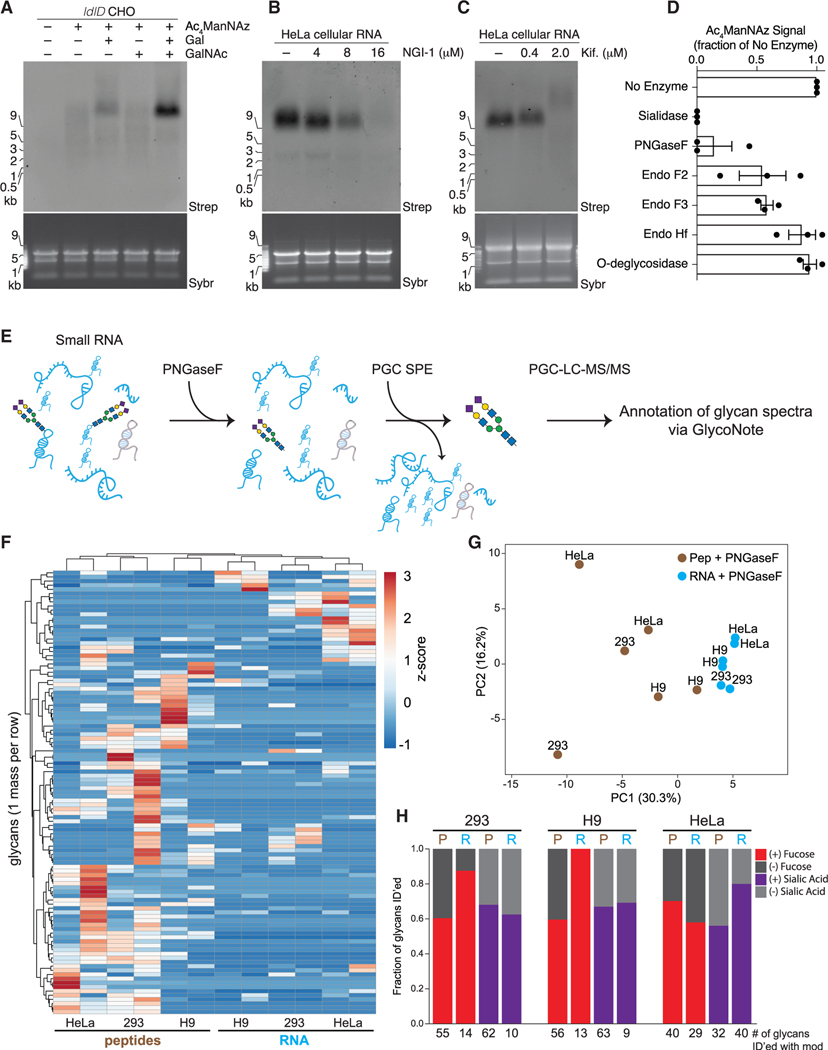
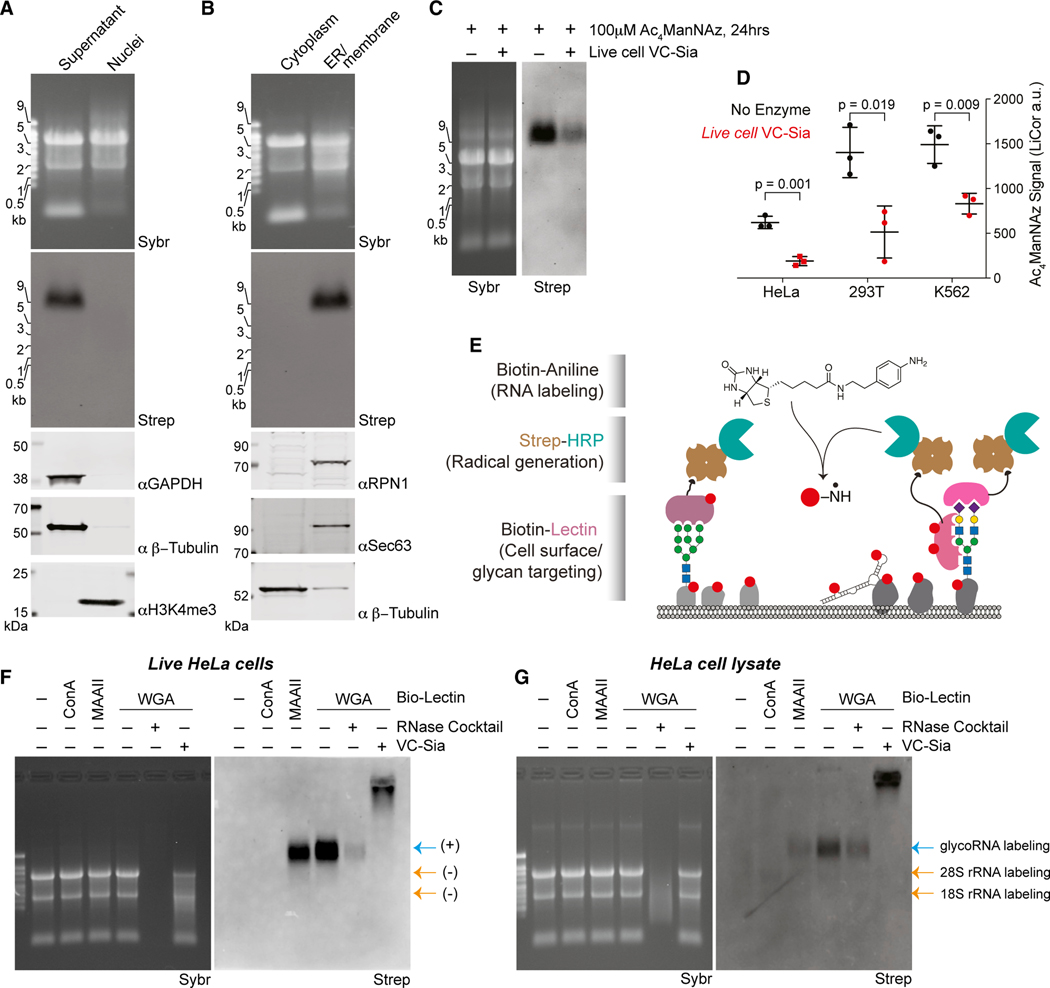
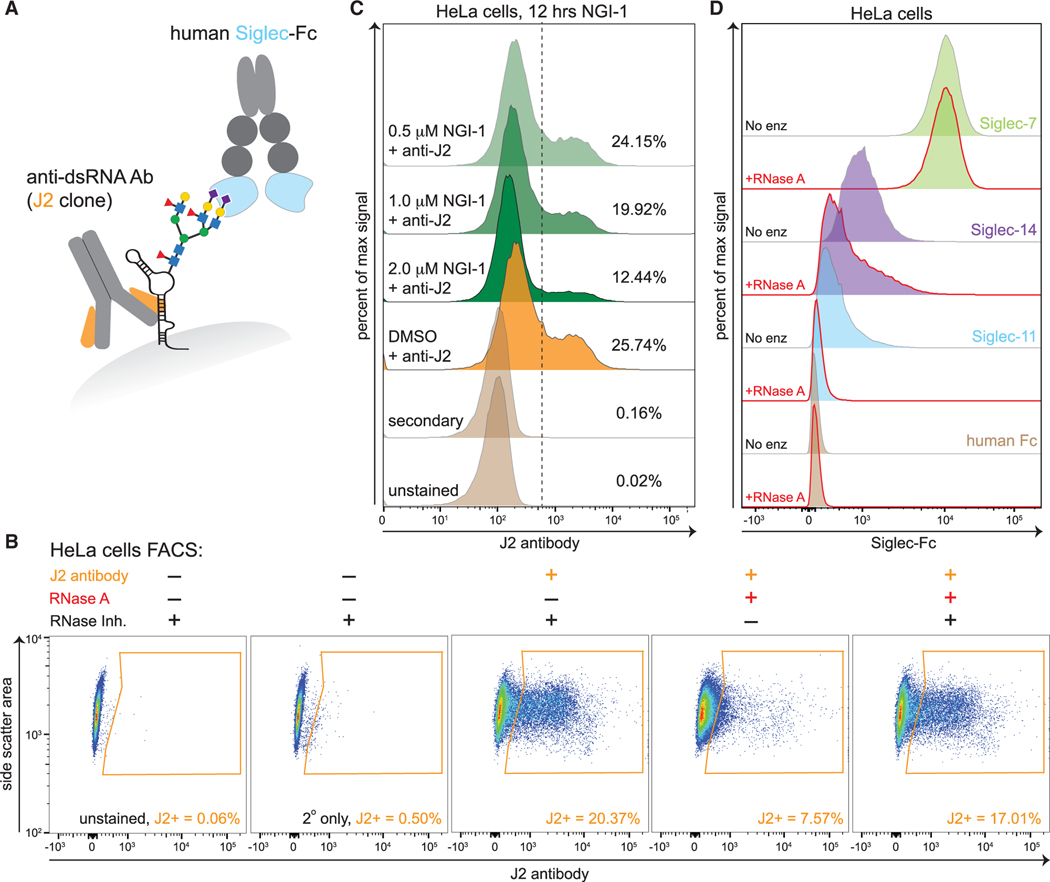
Glycans modify lipids and proteins to mediate inter- and intramolecular interactions across all domains of life. RNA is not thought to be a major target of glycosylation. Here, we challenge this view with evidence that mammals use RNA as a third scaffold for glycosylation. Using a battery of chemical and biochemical approaches, we found that conserved small noncoding RNAs bear sialylated glycans. These "glycoRNAs" were present in multiple cell types and mammalian species, in cultured cells, and in vivo. GlycoRNA assembly depends on canonical N-glycan biosynthetic machinery and results in structures enriched in sialic acid and fucose. Analysis of living cells revealed that the majority of glycoRNAs were present on the cell surface and can interact with anti-dsRNA antibodies and members of the Siglec receptor family. Collectively, these findings suggest the existence of a direct interface between RNA biology and glycobiology, and an expanded role for RNA in extracellular biology.
Keywords: RNA biology; glycoRNA; glycobiology; siglec; small RNAs.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests C.R.B. is co-founder and SAB member of Redwood Bioscience (a subsidiary of Catalent), Enable Biosciences, Palleon Pharmaceuticals, InterVenn Bio, OliLux Bio, and Lycia Therapeutics and a member of the Board of Directors of Eli Lilly.
Figures
Comment in
-
A glimpse at the glycoRNA world.Cell. 2021 Jun 10;184(12):3080-3081. doi: 10.1016/j.cell.2021.05.025. Cell. 2021. PMID: 34115968
-
Sugar-coated RNAs.Nat Rev Genet. 2021 Aug;22(8):480. doi: 10.1038/s41576-021-00388-y. Nat Rev Genet. 2021. PMID: 34168329 No abstract available.
-
RNA, a new member in the glycan-club that gets exposed at the cell surface.Traffic. 2021 Oct;22(10):362-363. doi: 10.1111/tra.12810. Epub 2021 Aug 17. Traffic. 2021. PMID: 34338403
References
-
- Agard NJ, Prescher JA, and Bertozzi CR (2004). A strain-promoted [3 + 2] azide-alkyne cycloaddition for covalent modification of biomolecules in living systems. J. Am. Chem. Soc 126, 15046–15047. - PubMed
-
- Ahlin E, Mathsson L, Eloranta M-L, Jonsdottir T, Gunnarsson I, Rönnblom L, and Rönnelid J. (2012). Autoantibodies associated with RNA are more enriched than anti-dsDNA antibodies in circulating immune complexes in SLE. Lupus 21, 586–595. - PubMed
-
- Angata T. (2014). Associations of genetic polymorphisms of Siglecs with human diseases. Glycobiology 24, 785–793. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
