

PPARγ-induced upregulation of subcutaneous fat adiponectin secretion, glyceroneogenesis and BCAA oxidation requires mTORC1 activity
- PMID: 34004356
- PMCID: PMC9391032
- DOI: 10.1016/j.bbalip.2021.158967
PPARγ-induced upregulation of subcutaneous fat adiponectin secretion, glyceroneogenesis and BCAA oxidation requires mTORC1 activity
Abstract
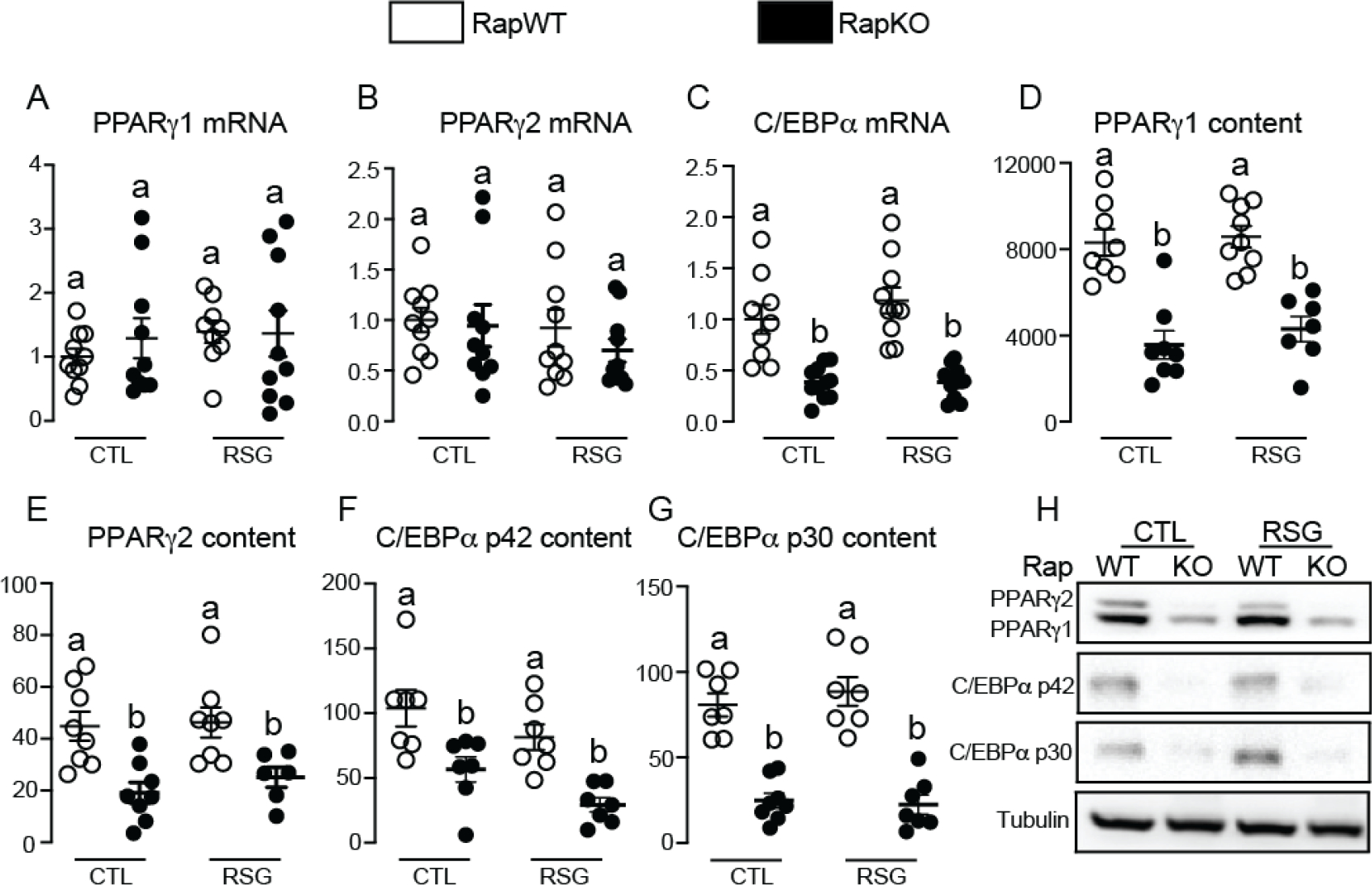
The nutrient sensors peroxisome proliferator-activated receptor γ (PPARγ) and mechanistic target of rapamycin complex 1 (mTORC1) closely interact in the regulation of adipocyte lipid storage. The precise mechanisms underlying this interaction and whether this extends to other metabolic processes and the endocrine function of adipocytes are still unknown. We investigated herein the involvement of mTORC1 as a mediator of the actions of the PPARγ ligand rosiglitazone in subcutaneous inguinal white adipose tissue (iWAT) mass, endocrine function, lipidome, transcriptome and branched-chain amino acid (BCAA) metabolism. Mice bearing regulatory associated protein of mTOR (Raptor) deletion and therefore mTORC1 deficiency exclusively in adipocytes and littermate controls were fed a high-fat diet supplemented or not with the PPARγ agonist rosiglitazone (30 mg/kg/day) for 8 weeks and evaluated for iWAT mass, lipidome, transcriptome (Rnaseq), respiration and BCAA metabolism. Adipocyte mTORC1 deficiency not only impaired iWAT adiponectin transcription, synthesis and secretion, PEPCK mRNA levels, triacylglycerol synthesis and BCAA oxidation and mRNA levels of related proteins but also completely blocked the upregulation in these processes induced by pharmacological PPARγ activation with rosiglitazone. Mechanistically, adipocyte mTORC1 deficiency impairs PPARγ transcriptional activity by reducing PPARγ protein content, as well as by downregulating C/EBPα, a co-partner and facilitator of PPARγ. In conclusion, mTORC1 and PPARγ are essential partners involved in the regulation of subcutaneous adipose tissue adiponectin production and secretion and BCAA oxidative metabolism.
Keywords: Adiponectin secretion; BCAA oxidation; C/EBPα; Glyceroneogenesis; PPARγ; Subcutaneous adipose tissue; mTORC1.
Copyright © 2021 Elsevier B.V. All rights reserved.
Conflict of interest statement
Conflict of Interests
Authors do not have any conflict of interest relevant to this article.
Figures
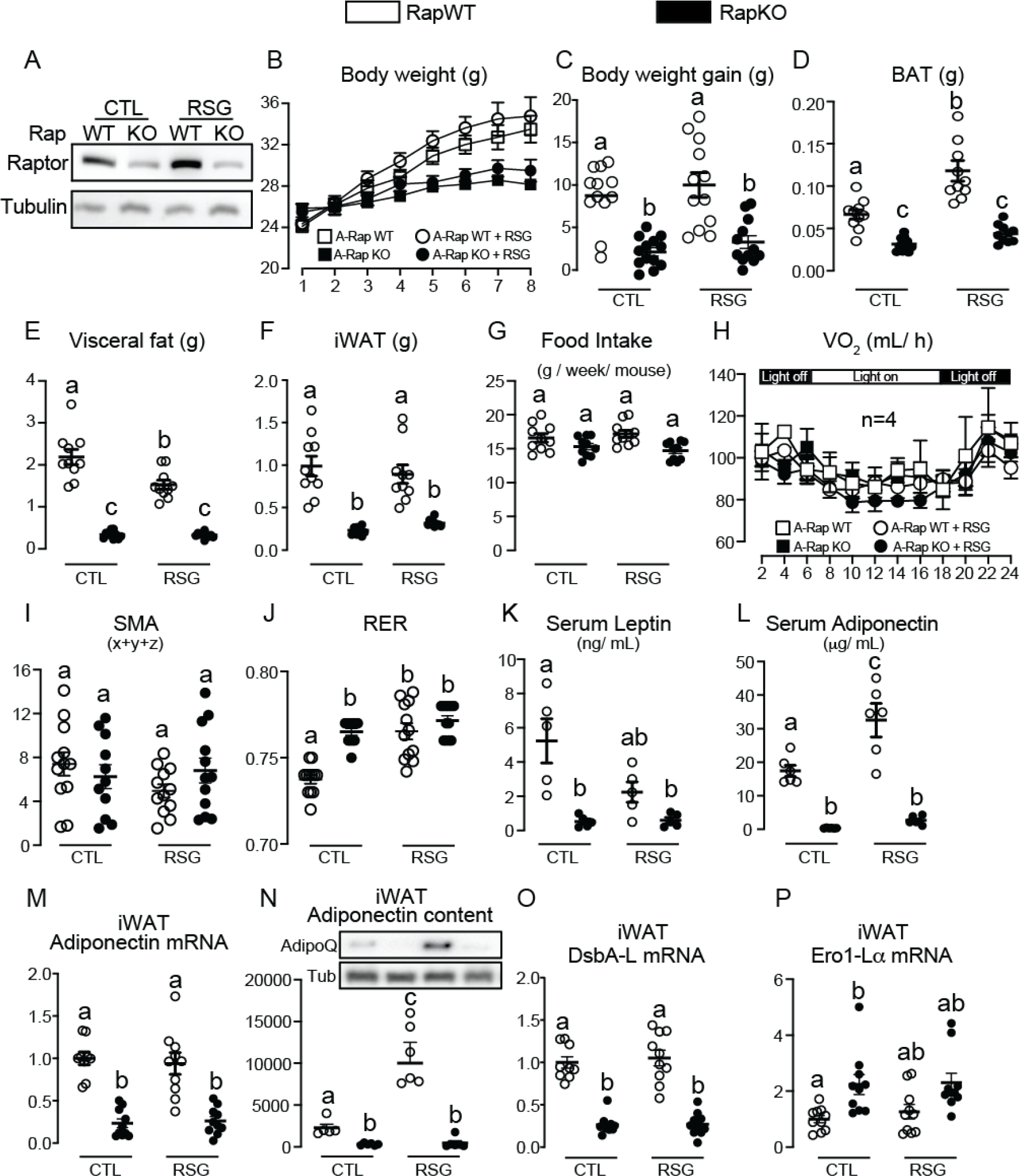
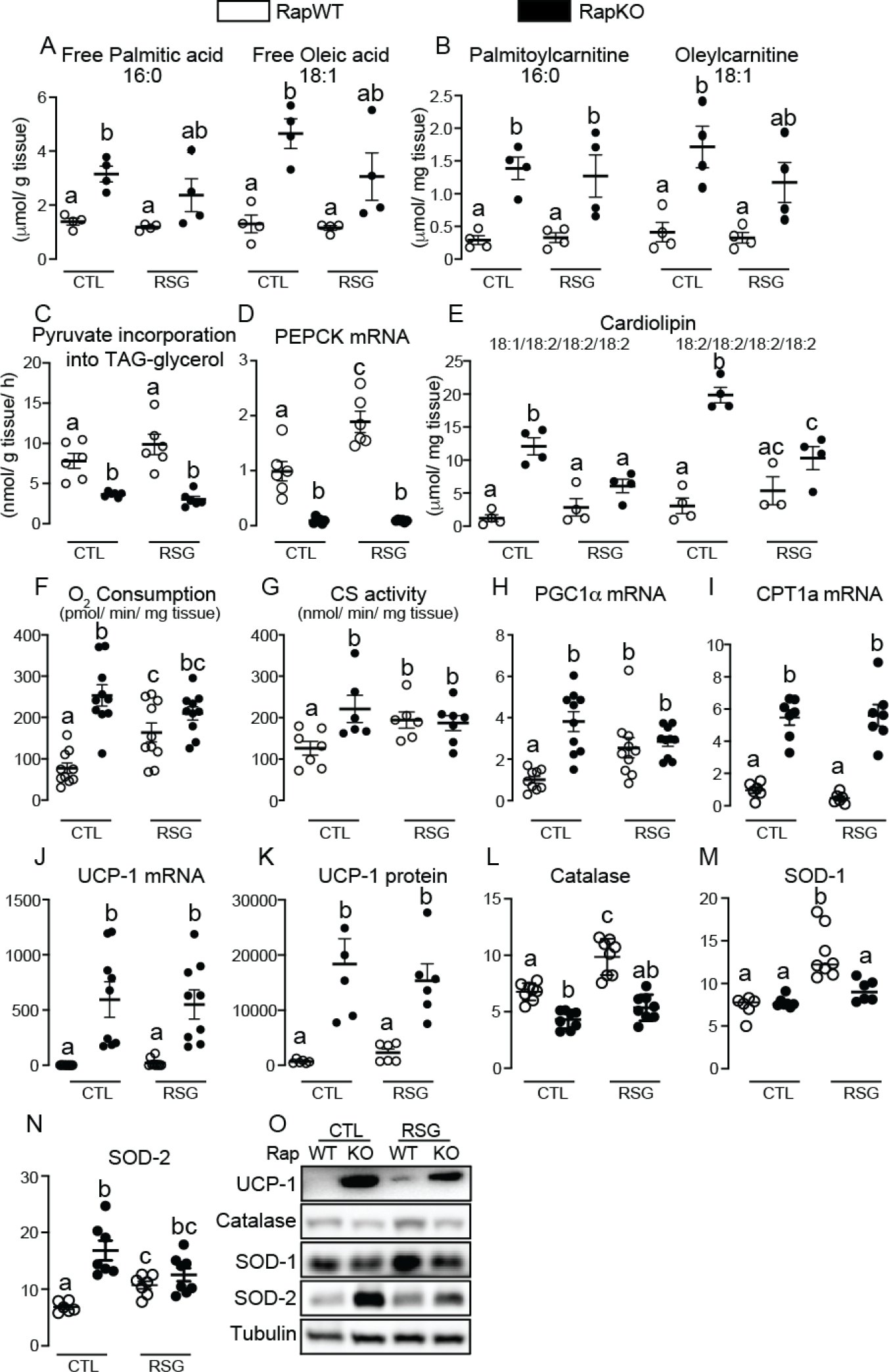
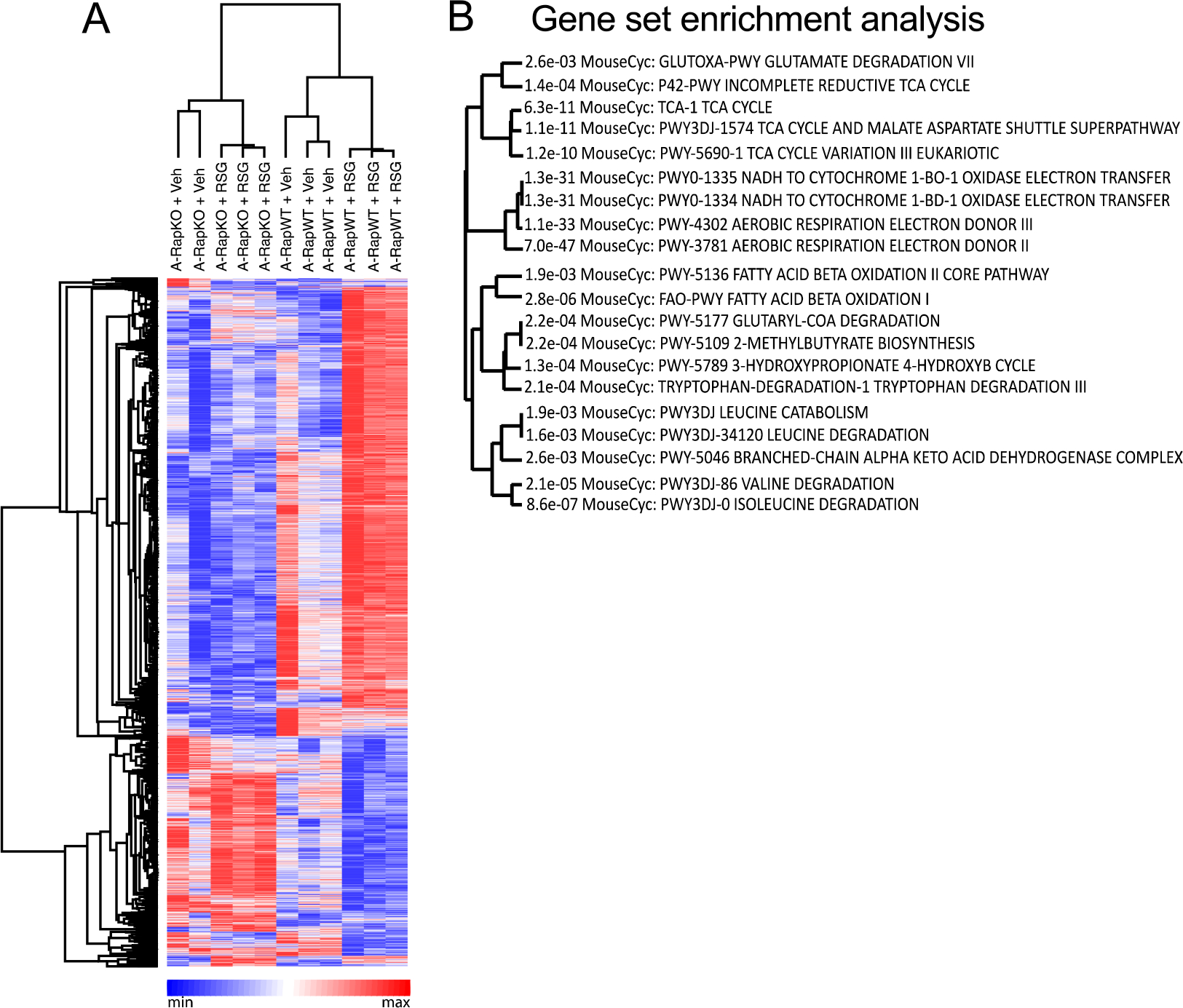
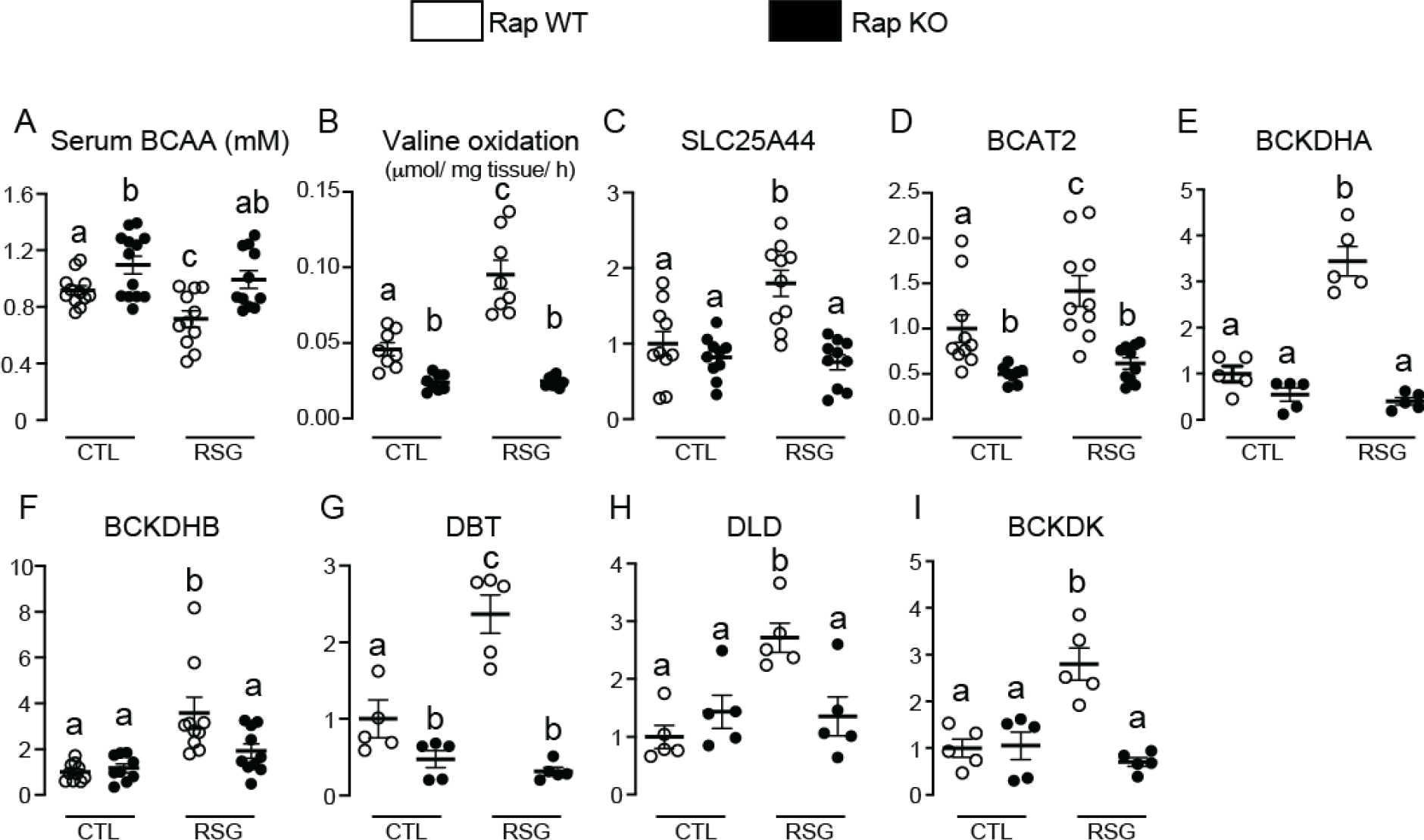
References
-
- Barroso I, Gurnell M, Crowley VE, Agostini M, Schwabe JW, Soos MA, et al., 1999. Dominant negative mutations in human PPARgamma associated with severe insulin resistance, diabetes mellitus and hypertension. Nature 402(6764):880–883. - PubMed
-
- Moreira RJ, Castro É, Oliveira TE, Belchior T, Peixoto AS, Chaves-Filho AB, et al., 2020. Lipoatrophy-Associated Insulin Resistance and Hepatic Steatosis are Attenuated by Intake of Diet Rich in Omega 3 Fatty Acids. Mol Nutr Food Res:e1900833. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
