

Neuromodulation and neuroprotective effects of chlorogenic acids in excitatory synapses of mouse hippocampal slices
- PMID: 34006978
- PMCID: PMC8131611
- DOI: 10.1038/s41598-021-89964-0
Neuromodulation and neuroprotective effects of chlorogenic acids in excitatory synapses of mouse hippocampal slices
Abstract
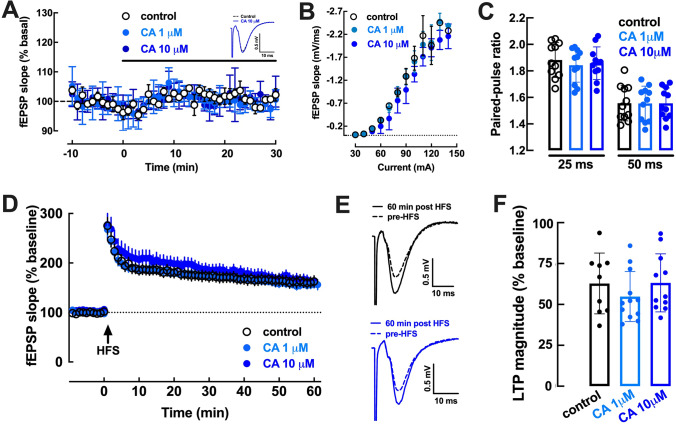
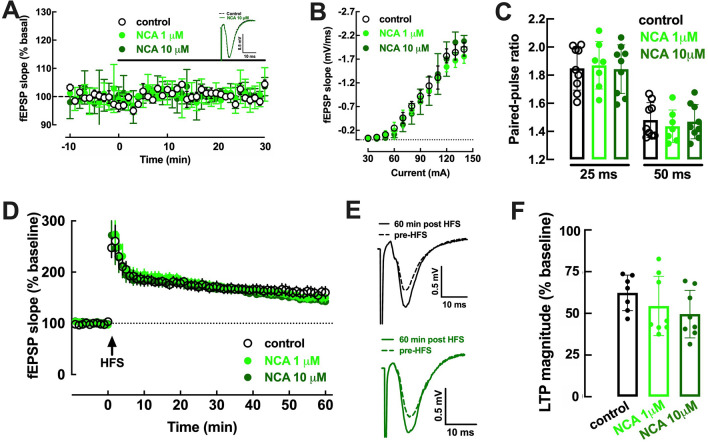
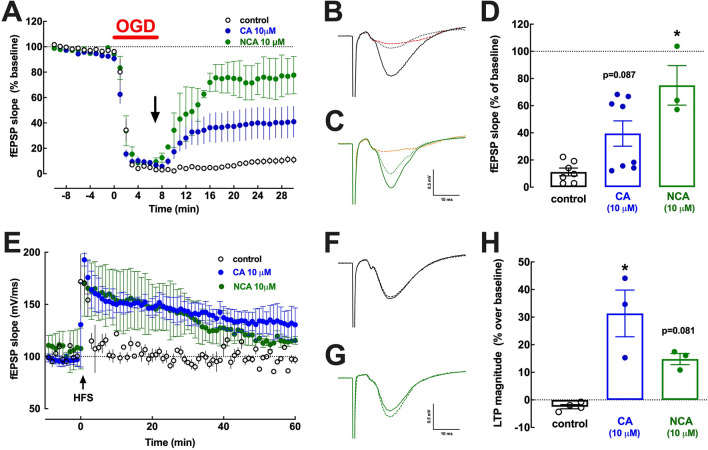
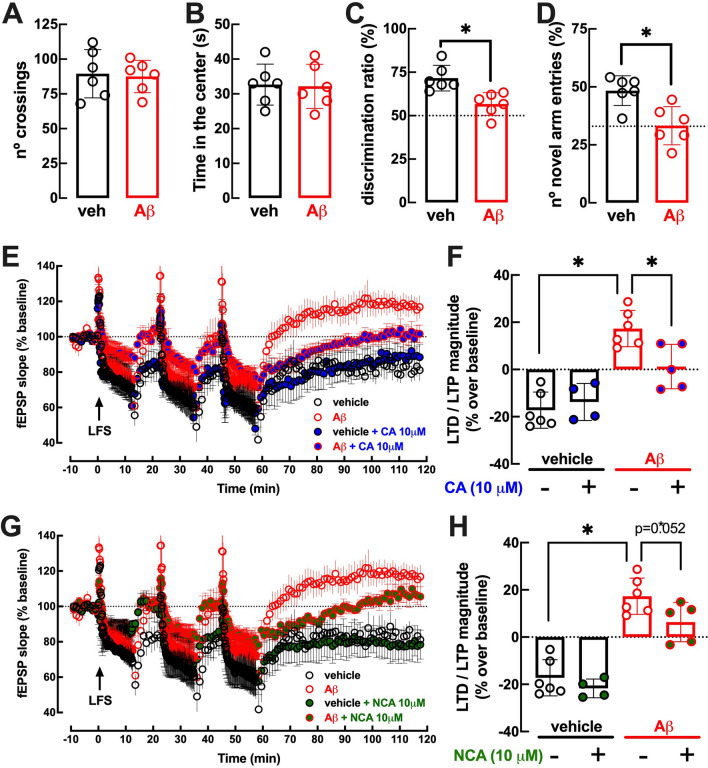
The increased healthspan afforded by coffee intake provides novel opportunities to identify new therapeutic strategies. Caffeine has been proposed to afford benefits through adenosine A2A receptors, which can control synaptic dysfunction underlying some brain disease. However, decaffeinated coffee and other main components of coffee such as chlorogenic acids, also attenuate brain dysfunction, although it is unknown if they control synaptic function. We now used electrophysiological recordings in mouse hippocampal slices to test if realistic concentrations of chlorogenic acids directly affect synaptic transmission and plasticity. 3-(3,4-dihydroxycinnamoyl)quinic acid (CA, 1-10 μM) and 5-O-(trans-3,4-dihydroxycinnamoyl)-D-quinic acid (NCA, 1-10 μM) were devoid of effect on synaptic transmission, paired-pulse facilitation or long-term potentiation (LTP) and long-term depression (LTD) in Schaffer collaterals-CA1 pyramidal synapses. However, CA and NCA increased the recovery of synaptic transmission upon re-oxygenation following 7 min of oxygen/glucose deprivation, an in vitro ischemia model. Also, CA and NCA attenuated the shift of LTD into LTP observed in hippocampal slices from animals with hippocampal-dependent memory deterioration after exposure to β-amyloid 1-42 (2 nmol, icv), in the context of Alzheimer's disease. These findings show that chlorogenic acids do not directly affect synaptic transmission and plasticity but can indirectly affect other cellular targets to correct synaptic dysfunction. Unraveling the molecular mechanisms of action of chlorogenic acids will allow the design of hitherto unrecognized novel neuroprotective strategies.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Caffeic acid recovers ischemia-induced synaptic dysfunction without direct effects on excitatory synaptic transmission and plasticity in mouse hippocampal slices.Neurosci Lett. 2023 Jun 21;808:137292. doi: 10.1016/j.neulet.2023.137292. Epub 2023 May 6. Neurosci Lett. 2023. PMID: 37156440
-
The physiological effects of caffeine on synaptic transmission and plasticity in the mouse hippocampus selectively depend on adenosine A1 and A2A receptors.Biochem Pharmacol. 2019 Aug;166:313-321. doi: 10.1016/j.bcp.2019.06.008. Epub 2019 Jun 12. Biochem Pharmacol. 2019. PMID: 31199895
-
Clinically relevant concentrations of ketamine mainly affect long-term potentiation rather than basal excitatory synaptic transmission and do not change paired-pulse facilitation in mouse hippocampal slices.Brain Res. 2014 Apr 29;1560:10-7. doi: 10.1016/j.brainres.2014.03.004. Epub 2014 Mar 15. Brain Res. 2014. PMID: 24637258
-
Synaptic modulation by coffee compounds: Insights into neural plasticity.Prog Brain Res. 2024;289:181-191. doi: 10.1016/bs.pbr.2024.06.008. Epub 2024 Jun 21. Prog Brain Res. 2024. PMID: 39168580 Review.
-
Digging Deeper: Advancements in Visualization of Inhibitory Synapses in Neurodegenerative Disorders.Int J Mol Sci. 2021 Nov 18;22(22):12470. doi: 10.3390/ijms222212470. Int J Mol Sci. 2021. PMID: 34830352 Free PMC article. Review.
Cited by
-
Identification of a Hydroxygallic Acid Derivative, Zingibroside R1 and a Sterol Lipid as Potential Active Ingredients of Cuscuta chinensis Extract That Has Neuroprotective and Antioxidant Effects in Aged Caenorhabditis elegans.Nutrients. 2022 Oct 9;14(19):4199. doi: 10.3390/nu14194199. Nutrients. 2022. PMID: 36235851 Free PMC article.
-
Chlorogenic Acid Decreases Glutamate Release from Rat Cortical Nerve Terminals by P/Q-Type Ca2+ Channel Suppression: A Possible Neuroprotective Mechanism.Int J Mol Sci. 2021 Oct 23;22(21):11447. doi: 10.3390/ijms222111447. Int J Mol Sci. 2021. PMID: 34768876 Free PMC article.
-
Review of Plant Extracts and Active Components: Mechanisms of Action for the Treatment of Obesity-Induced Cognitive Impairment.Brain Sci. 2023 Jun 8;13(6):929. doi: 10.3390/brainsci13060929. Brain Sci. 2023. PMID: 37371407 Free PMC article. Review.
-
Risk Assessment of Chlorogenic and Isochlorogenic Acids in Coffee By-Products.Molecules. 2023 Jul 20;28(14):5540. doi: 10.3390/molecules28145540. Molecules. 2023. PMID: 37513412 Free PMC article. Review.
-
Anti-Alzheimer's Potency of Rich Phenylethanoid Glycosides Extract from Marrubium vulgare L.: In Vitro and In Silico Studies.Pharmaceuticals (Basel). 2024 Sep 27;17(10):1282. doi: 10.3390/ph17101282. Pharmaceuticals (Basel). 2024. PMID: 39458923 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
