

JunB is a key regulator of multiple myeloma bone marrow angiogenesis
- PMID: 34007044
- PMCID: PMC8632680
- DOI: 10.1038/s41375-021-01271-9
JunB is a key regulator of multiple myeloma bone marrow angiogenesis
Erratum in
-
Publisher Correction: JunB is a key regulator of multiple myeloma bone marrow angiogenesis.Leukemia. 2021 Dec;35(12):3628. doi: 10.1038/s41375-021-01367-2. Leukemia. 2021. PMID: 34489554 Free PMC article. No abstract available.
Abstract
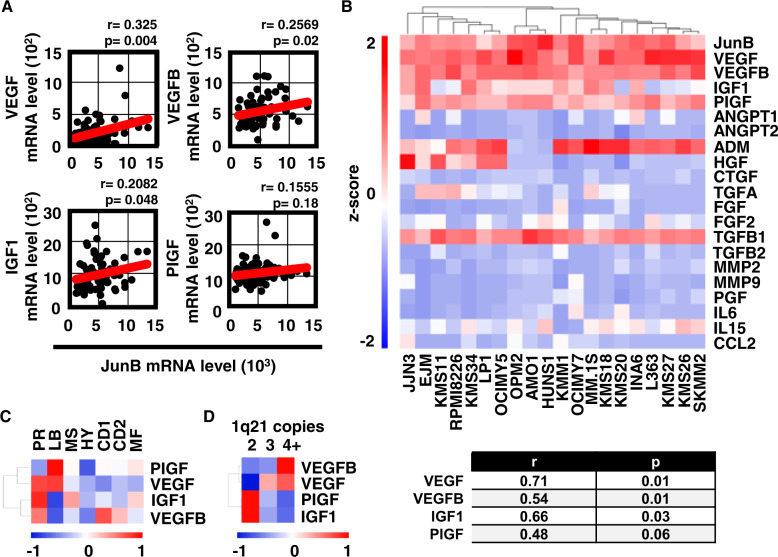
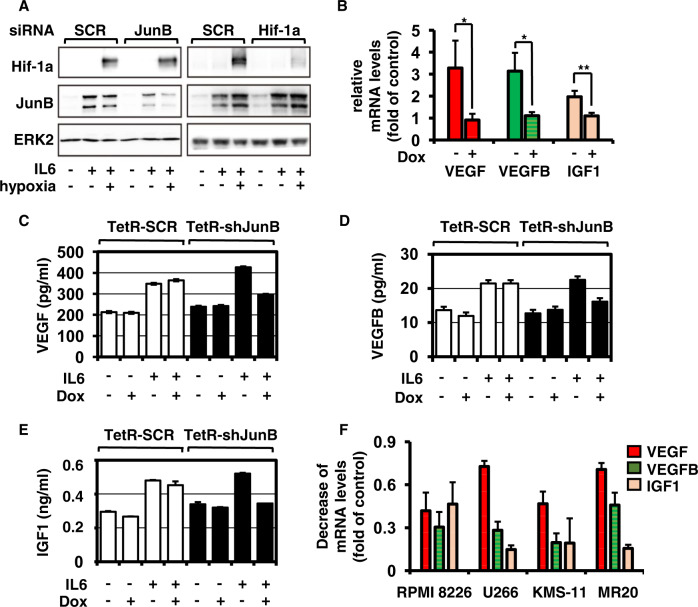
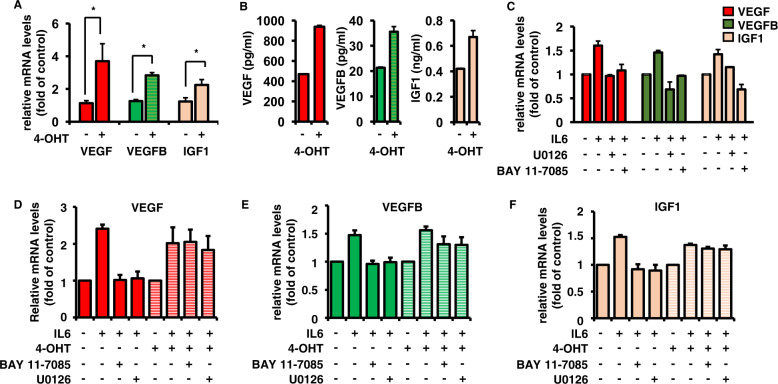
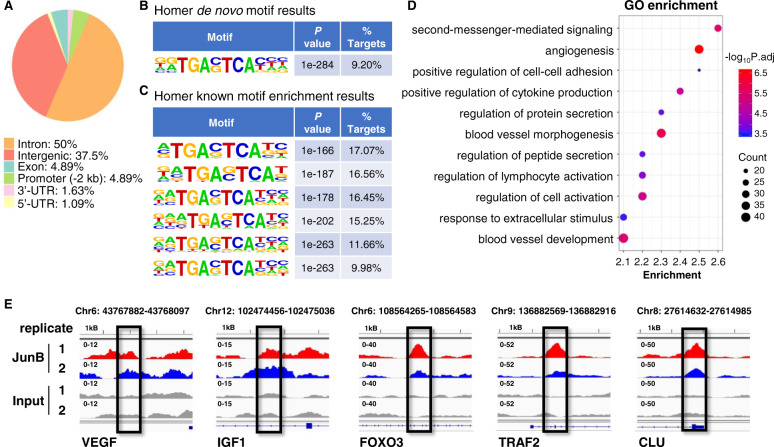
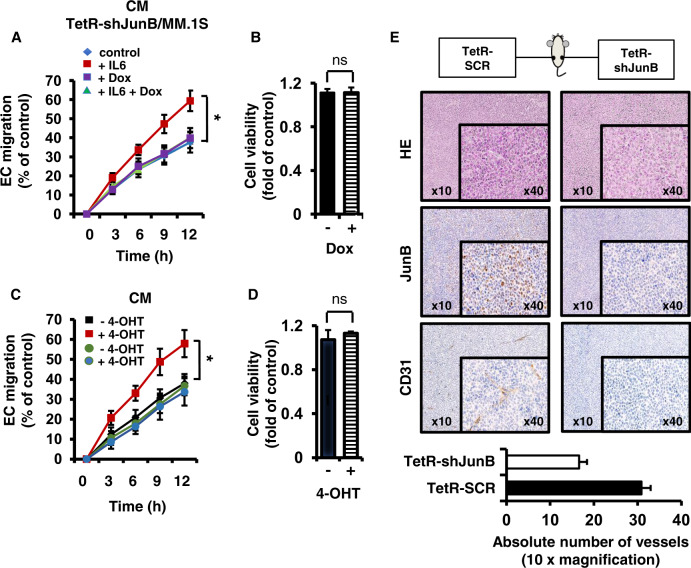
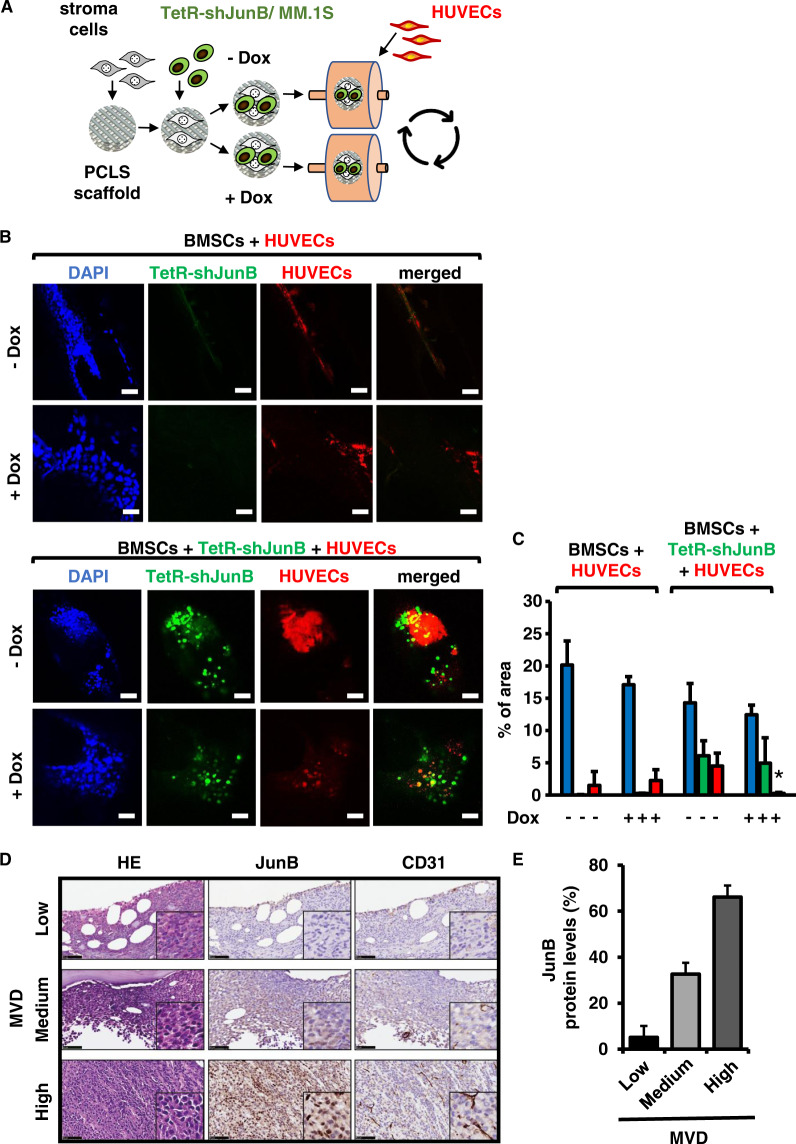
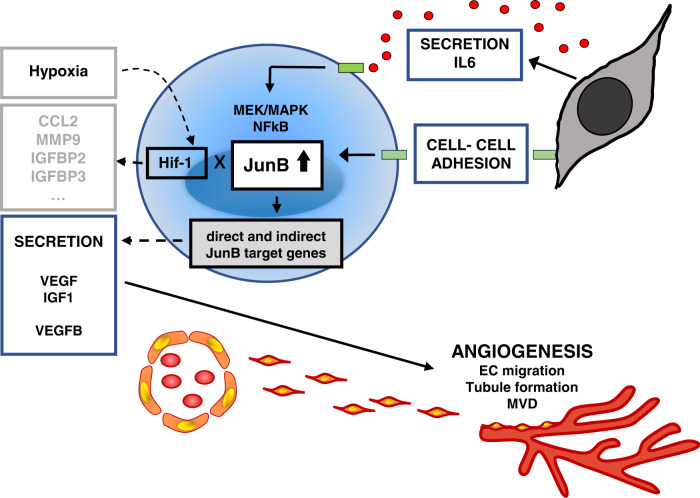
Bone marrow (BM) angiogenesis significantly influences disease progression in multiple myeloma (MM) patients and correlates with adverse prognosis. The present study shows a statistically significant correlation of the AP-1 family member JunB with VEGF, VEGFB, and IGF1 expression levels in MM. In contrast to the angiogenic master regulator Hif-1α, JunB protein levels were independent of hypoxia. Results in tumor-cell models that allow the induction of JunB knockdown or JunB activation, respectively, corroborated the functional role of JunB in the production and secretion of these angiogenic factors (AFs). Consequently, conditioned media derived from MM cells after JunB knockdown or JunB activation either inhibited or stimulated in vitro angiogenesis. The impact of JunB on MM BM angiogenesis was finally confirmed in a dynamic 3D model of the BM microenvironment, a xenograft mouse model as well as in patient-derived BM sections. In summary, in continuation of our previous study (Fan et al., 2017), the present report reveals for the first time that JunB is not only a mediator of MM cell survival, proliferation, and drug resistance, but also a promoter of AF transcription and consequently of MM BM angiogenesis. Our results thereby underscore worldwide efforts to target AP-1 transcription factors such as JunB as a promising strategy in MM therapy.
© 2021. The Author(s).
Conflict of interest statement
SV received speaker’s honoraria from Bristol Myers Squibb, MSD, Pfizer, and consultancy fees from Roche, Eusa, MSD, and Merck. DJ received consultant honoraria from Bayer, Amgen, MSD, CureVac, Roche, and BMS. KP has received speaker’s honoraria from Celgene, Amgen Inc., and Janssen Pharmaceuticals, consultancy fees from Celgene, Takeda, and Janssen Pharmaceuticals, and research support from Roche Pharmaceuticals. The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript apart from those disclosed. The remaining authors declare no conflict of interest.
Figures
References
-
- Vacca A, Ribatti D. Bone marrow angiogenesis in multiple myeloma. Leukemia. 2006;20:193–9. - PubMed
-
- Vacca A, Ribatti D, Roncali L, Ranieri G, Serio G, Silvestris F, et al. Bone marrow angiogenesis and progression in multiple myeloma. Br J Haematol. 1994;87:503–8. - PubMed
-
- Vacca A, Ria R, Semeraro F, Merchionne F, Coluccia M, Boccarelli A, et al. Endothelial cells in the bone marrow of patients with multiple myeloma. Blood. 2003;102:3340–8. - PubMed
-
- Nosàs-Garcia S, Moehler T, Wasser K, Kiessling F, Bartl R, Zuna I, et al. Dynamic contrast-enhanced MRI for assessing the disease activity of multiple myeloma: a comparative study with histology and clinical markers. J Magn Reson Imaging. 2005;22:154–62. - PubMed
-
- Vacca A, Ribatti D, Presta M, Minischetti M, Iurlaro M, Ria R, et al. Bone marrow neovascularization, plasma cell angiogenic potential, and matrix metalloproteinase-2 secretion parallel progression of human multiple myeloma. Blood. 1999;93:3064–73. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
