

Genomic evolution of the class Acidithiobacillia: deep-branching Proteobacteria living in extreme acidic conditions
- PMID: 34007059
- PMCID: PMC8528912
- DOI: 10.1038/s41396-021-00995-x
Genomic evolution of the class Acidithiobacillia: deep-branching Proteobacteria living in extreme acidic conditions
Abstract
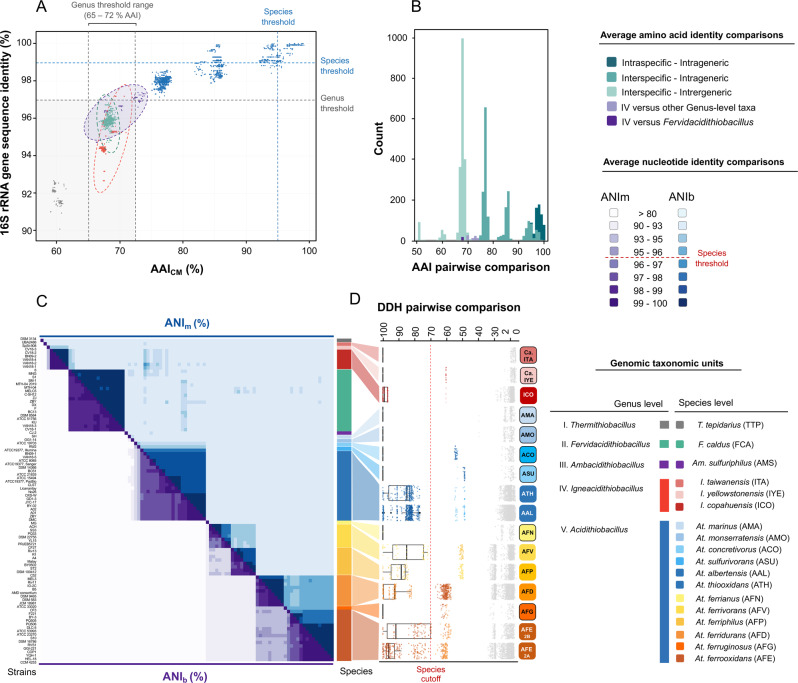
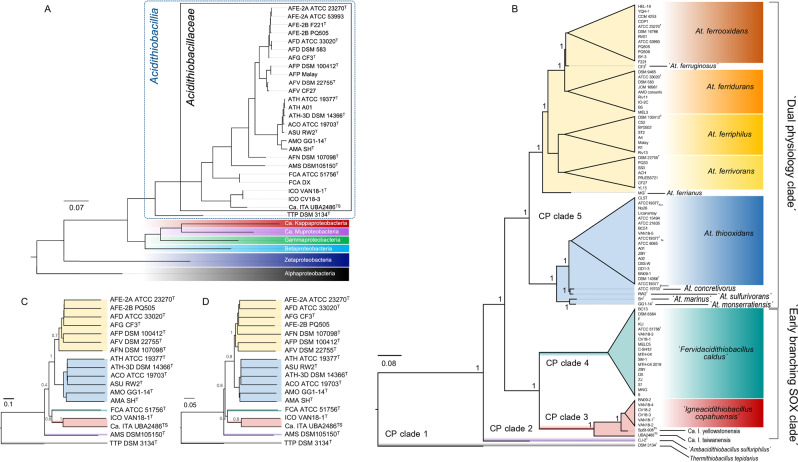
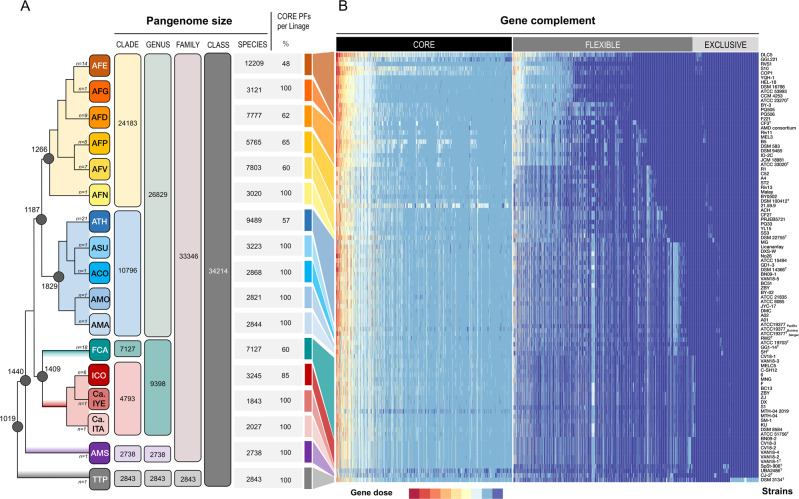
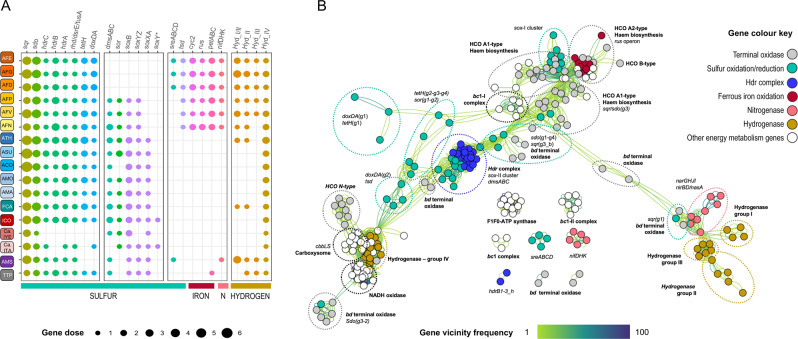
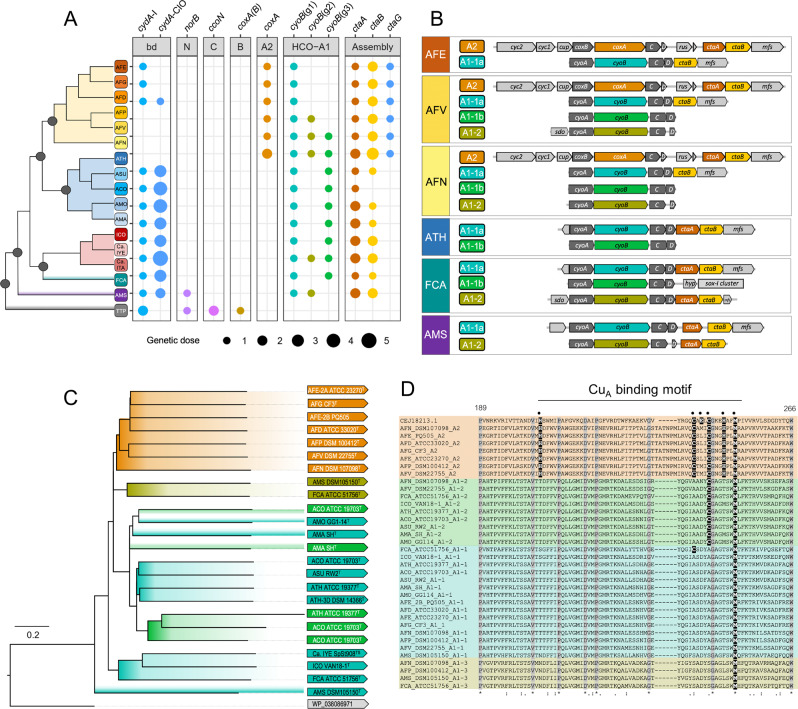
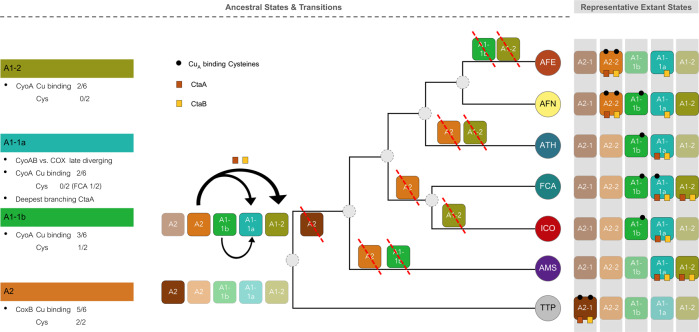
Members of the genus Acidithiobacillus, now ranked within the class Acidithiobacillia, are model bacteria for the study of chemolithotrophic energy conversion under extreme conditions. Knowledge of the genomic and taxonomic diversity of Acidithiobacillia is still limited. Here, we present a systematic analysis of nearly 100 genomes from the class sampled from a wide range of habitats. Some of these genomes are new and others have been reclassified on the basis of advanced genomic analysis, thus defining 19 Acidithiobacillia lineages ranking at different taxonomic levels. This work provides the most comprehensive classification and pangenomic analysis of this deep-branching class of Proteobacteria to date. The phylogenomic framework obtained illuminates not only the evolutionary past of this lineage, but also the molecular evolution of relevant aerobic respiratory proteins, namely the cytochrome bo3 ubiquinol oxidases.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Integrative Genomics Sheds Light on Evolutionary Forces Shaping the Acidithiobacillia Class Acidophilic Lifestyle.Front Microbiol. 2022 Feb 15;12:822229. doi: 10.3389/fmicb.2021.822229. eCollection 2021. Front Microbiol. 2022. PMID: 35242113 Free PMC article.
-
Molecular evolution of cytochrome bd oxidases across proteobacterial genomes.Genome Biol Evol. 2015 Feb 16;7(3):801-20. doi: 10.1093/gbe/evv032. Genome Biol Evol. 2015. PMID: 25688108 Free PMC article.
-
Comparative genomics sheds light on transcription factor-mediated regulation in the extreme acidophilic Acidithiobacillia representatives.Res Microbiol. 2024 Jan-Feb;175(1-2):104135. doi: 10.1016/j.resmic.2023.104135. Epub 2023 Sep 9. Res Microbiol. 2024. PMID: 37678513
-
The branching order and phylogenetic placement of species from completed bacterial genomes, based on conserved indels found in various proteins.Int Microbiol. 2001 Dec;4(4):187-202. doi: 10.1007/s10123-001-0037-9. Int Microbiol. 2001. PMID: 12051562 Review.
-
Challenges in gene-oriented approaches for pangenome content discovery.Brief Bioinform. 2021 May 20;22(3):bbaa198. doi: 10.1093/bib/bbaa198. Brief Bioinform. 2021. PMID: 32893299 Review.
Cited by
-
Integrative Genomics Sheds Light on Evolutionary Forces Shaping the Acidithiobacillia Class Acidophilic Lifestyle.Front Microbiol. 2022 Feb 15;12:822229. doi: 10.3389/fmicb.2021.822229. eCollection 2021. Front Microbiol. 2022. PMID: 35242113 Free PMC article.
-
AI-driven pan-proteome analyses reveal insights into the biohydrometallurgical properties of Acidithiobacillia.Front Microbiol. 2023 Sep 7;14:1243987. doi: 10.3389/fmicb.2023.1243987. eCollection 2023. Front Microbiol. 2023. PMID: 37744906 Free PMC article.
-
Evolution of Type IV CRISPR-Cas Systems: Insights from CRISPR Loci in Integrative Conjugative Elements of Acidithiobacillia.CRISPR J. 2021 Oct;4(5):656-672. doi: 10.1089/crispr.2021.0051. Epub 2021 Sep 28. CRISPR J. 2021. PMID: 34582696 Free PMC article.
-
QS molecules change the planktonic/mineral subpopulations distribution of moderately thermophilic leaching bacteria in pyrite and decrease leaching in chalcopyrite.Front Microbiol. 2025 May 16;16:1592588. doi: 10.3389/fmicb.2025.1592588. eCollection 2025. Front Microbiol. 2025. PMID: 40454366 Free PMC article.
-
Molecular Identification and Acid Stress Response of an Acidithiobacillus thiooxidans Strain Isolated from Rio Tinto (Spain).Int J Mol Sci. 2023 Aug 29;24(17):13391. doi: 10.3390/ijms241713391. Int J Mol Sci. 2023. PMID: 37686204 Free PMC article.
References
-
- Boden R, Hutt LP. Acidithiobacillus. In Bergey’s manual of systematics of archaea and bacteria. Wiley; 2019.
-
- Johnson DB. The evolution, current status, and future prospects of using biotechnologies in the mineral extraction and metal recovery sectors. Minerals. 2018;8:343. doi: 10.3390/min8080343. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
