

Onset of taste bud cell renewal starts at birth and coincides with a shift in SHH function
- PMID: 34009125
- PMCID: PMC8172241
- DOI: 10.7554/eLife.64013
Onset of taste bud cell renewal starts at birth and coincides with a shift in SHH function
Abstract
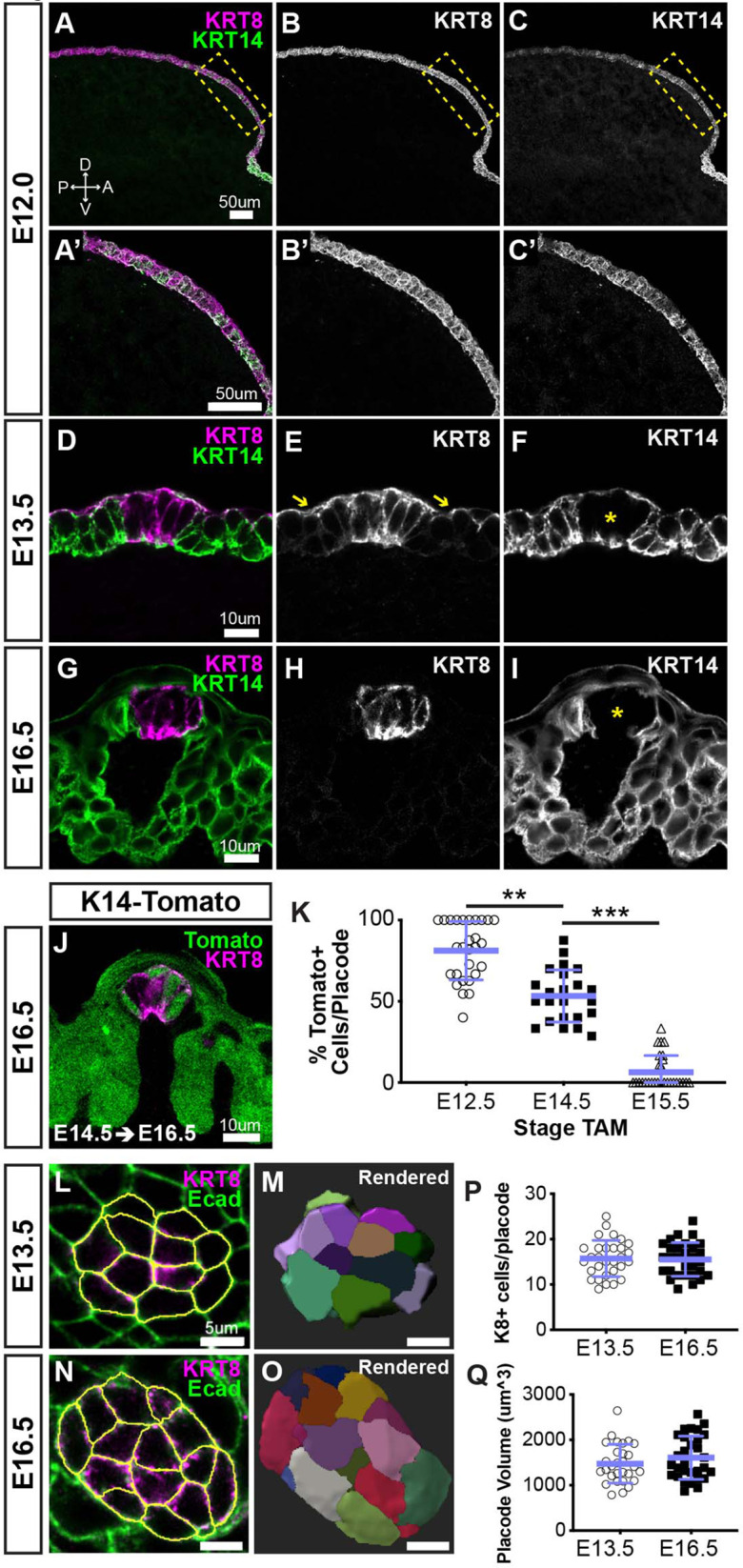
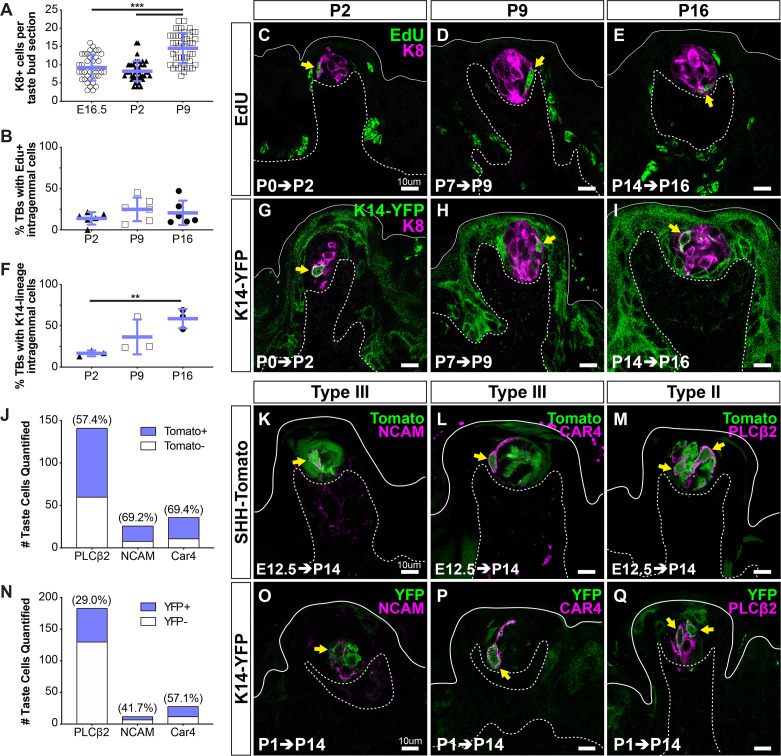
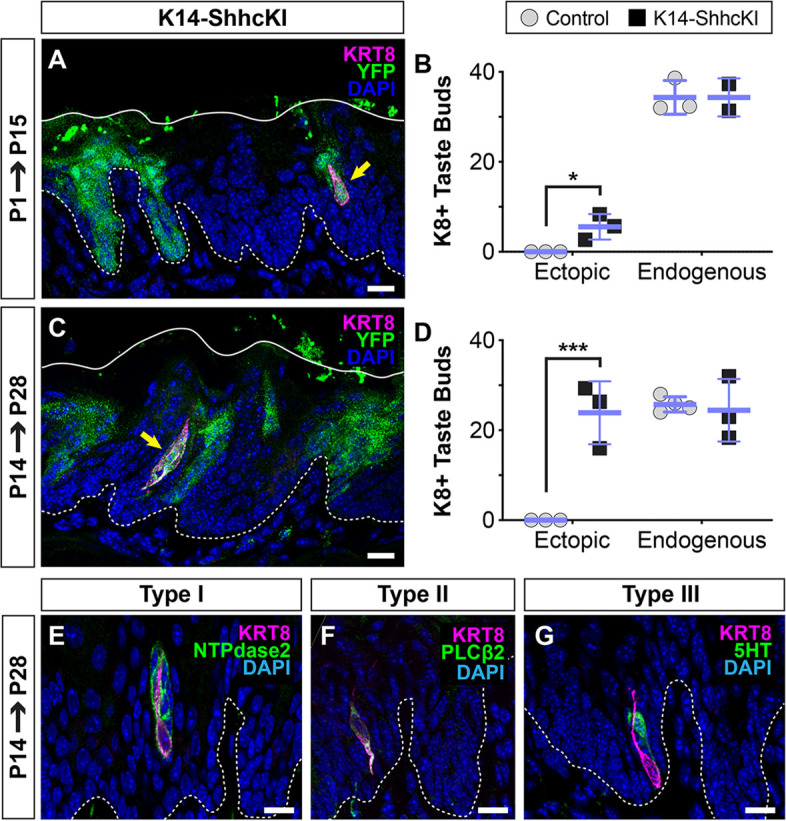
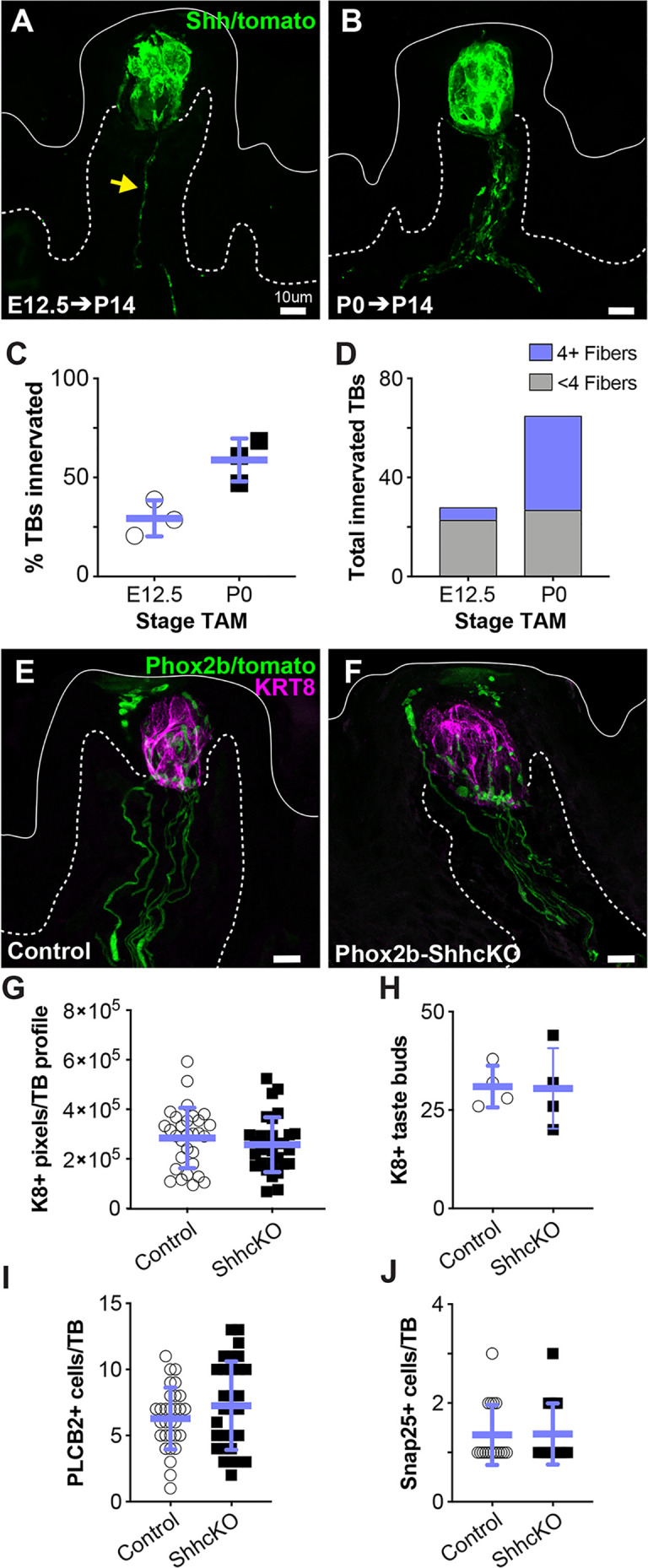
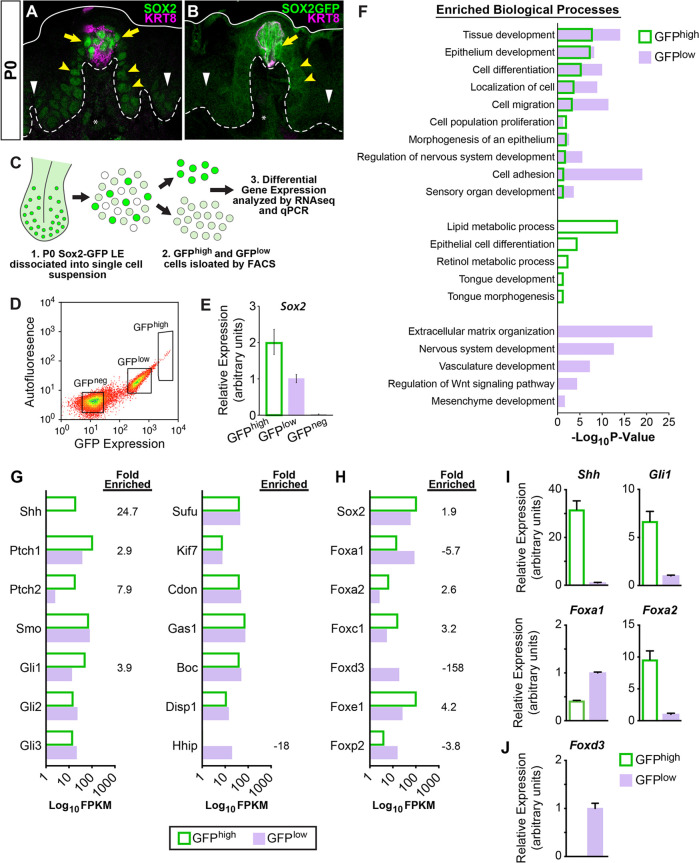
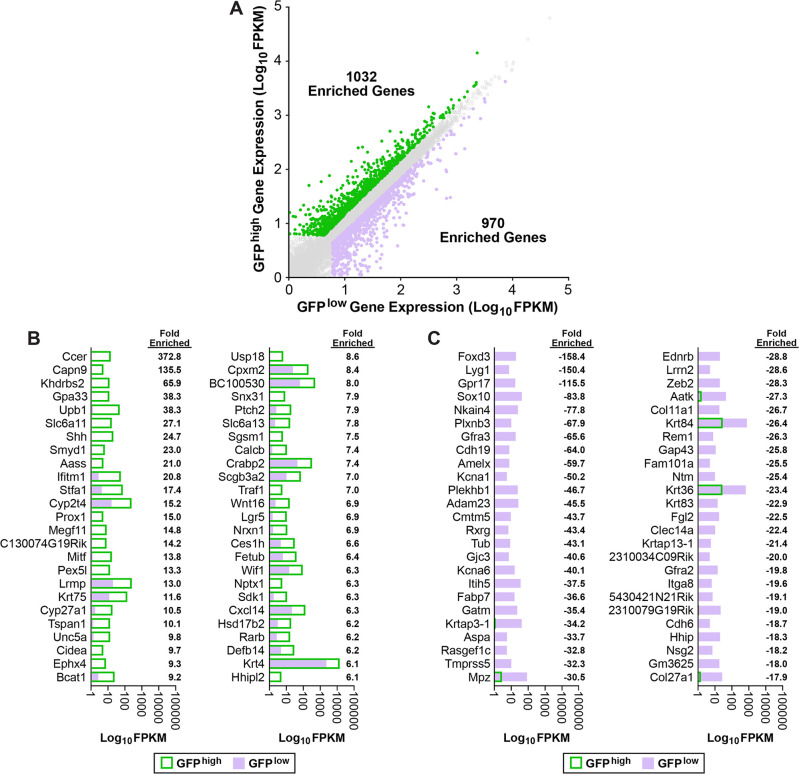
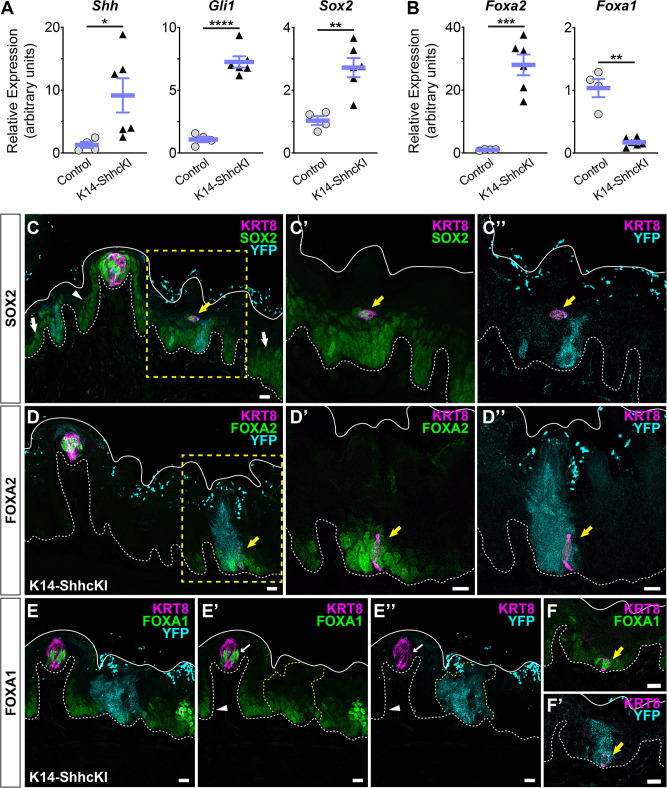
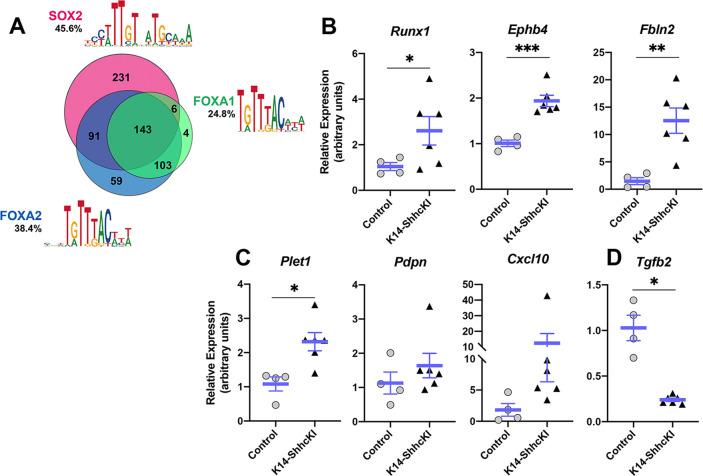
Embryonic taste bud primordia are specified as taste placodes on the tongue surface and differentiate into the first taste receptor cells (TRCs) at birth. Throughout adult life, TRCs are continually regenerated from epithelial progenitors. Sonic hedgehog (SHH) signaling regulates TRC development and renewal, repressing taste fate embryonically, but promoting TRC differentiation in adults. Here, using mouse models, we show TRC renewal initiates at birth and coincides with onset of SHHs pro-taste function. Using transcriptional profiling to explore molecular regulators of renewal, we identified Foxa1 and Foxa2 as potential SHH target genes in lingual progenitors at birth and show that SHH overexpression in vivo alters FoxA1 and FoxA2 expression relevant to taste buds. We further bioinformatically identify genes relevant to cell adhesion and cell locomotion likely regulated by FOXA1;FOXA2 and show that expression of these candidates is also altered by forced SHH expression. We present a new model where SHH promotes TRC differentiation by regulating changes in epithelial cell adhesion and migration.
Keywords: Hedgehog signaling; RNAseq; SOX2; developmental biology; mouse; regeneration; regenerative medicine; stem cells; taste; taste bud.
© 2021, Golden et al.
Conflict of interest statement
EG, EL, LS, GT, DG, TF, JS, KJ, LB No competing interests declared
Figures




References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
