

Per1/Per2-Igf2 axis-mediated circadian regulation of myogenic differentiation
- PMID: 34009269
- PMCID: PMC8138781
- DOI: 10.1083/jcb.202101057
Per1/Per2-Igf2 axis-mediated circadian regulation of myogenic differentiation
Abstract
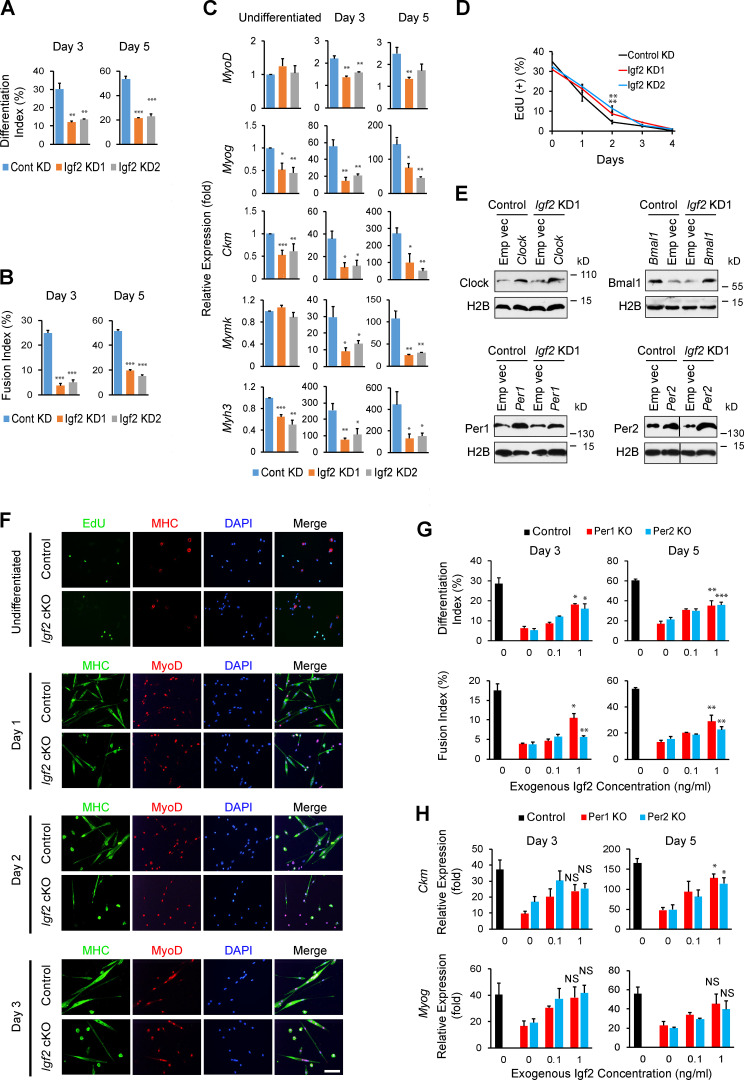
Circadian rhythms regulate cell proliferation and differentiation, but circadian control of tissue regeneration remains elusive at the molecular level. Here, we show that proper myoblast differentiation and muscle regeneration are regulated by the circadian master regulators Per1 and Per2. Depletion of Per1 or Per2 suppressed myoblast differentiation in vitro and muscle regeneration in vivo, demonstrating their nonredundant functions. Both Per1 and Per2 were required for the activation of Igf2, an autocrine promoter of myoblast differentiation, accompanied by Per-dependent recruitment of RNA polymerase II, dynamic histone modifications at the Igf2 promoter and enhancer, and the promoter-enhancer interaction. This circadian epigenetic priming created a preferred time window for initiating myoblast differentiation. Consistently, muscle regeneration was faster if initiated at night, when Per1, Per2, and Igf2 were highly expressed compared with morning. This study reveals the circadian timing as a significant factor for effective muscle cell differentiation and regeneration.
© 2021 Katoku-Kikyo et al.
Figures











References
-
- Andrews, J.L., Zhang X., McCarthy J.J., McDearmon E.L., Hornberger T.A., Russell B., Campbell K.S., Arbogast S., Reid M.B., Walker J.R., et al. . 2010. CLOCK and BMAL1 regulate MyoD and are necessary for maintenance of skeletal muscle phenotype and function. Proc. Natl. Acad. Sci. USA. 107:19090–19095. 10.1073/pnas.1014523107 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
