

Mechanistic model of nutrient uptake explains dichotomy between marine oligotrophic and copiotrophic bacteria
- PMID: 34010286
- PMCID: PMC8168909
- DOI: 10.1371/journal.pcbi.1009023
Mechanistic model of nutrient uptake explains dichotomy between marine oligotrophic and copiotrophic bacteria
Abstract
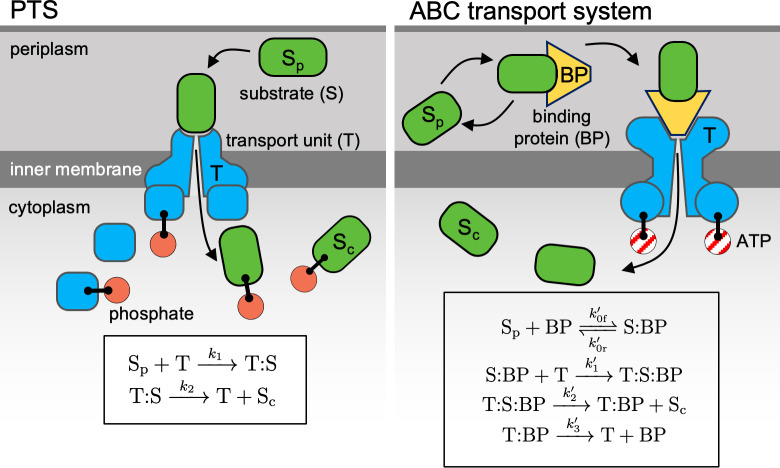
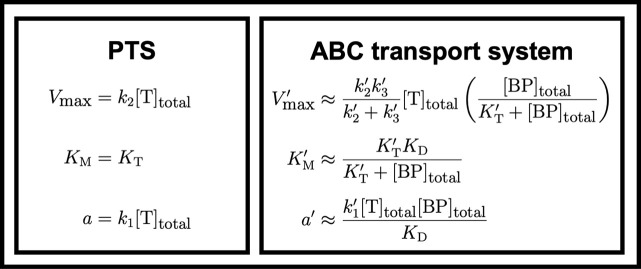
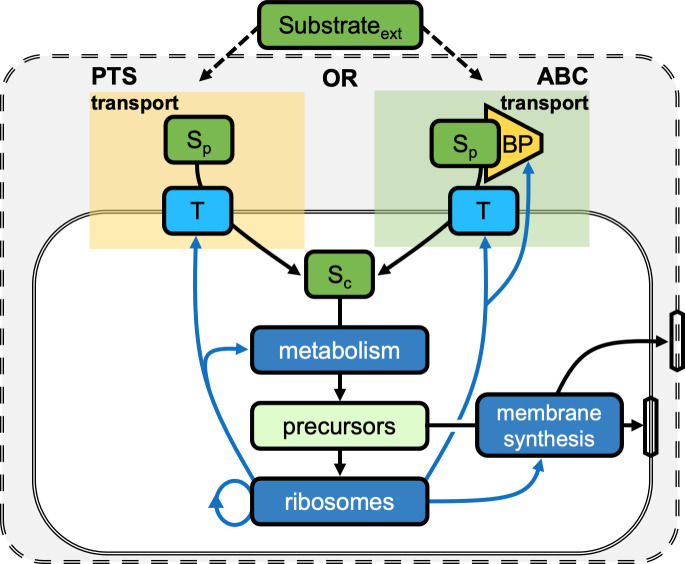
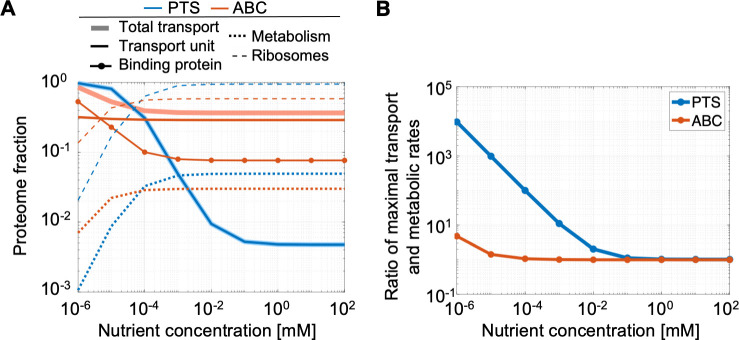
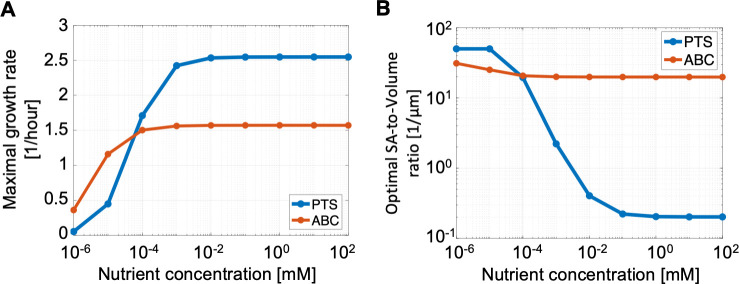
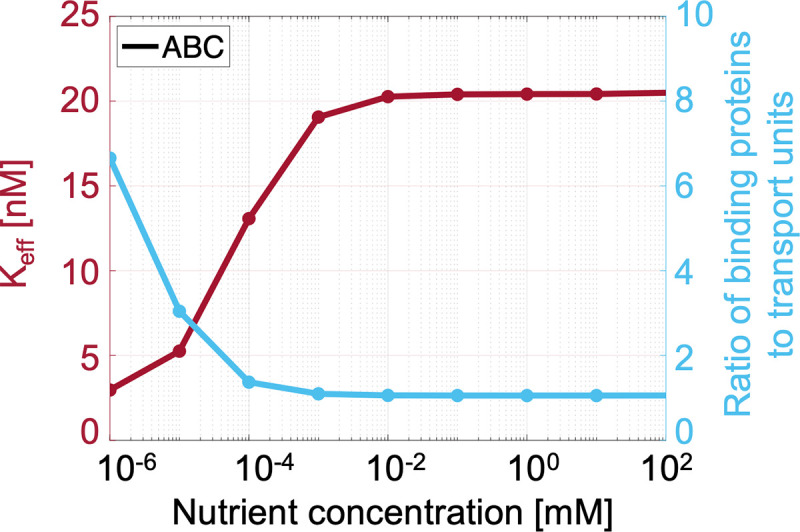
Marine bacterial diversity is immense and believed to be driven in part by trade-offs in metabolic strategies. Here we consider heterotrophs that rely on organic carbon as an energy source and present a molecular-level model of cell metabolism that explains the dichotomy between copiotrophs-which dominate in carbon-rich environments-and oligotrophs-which dominate in carbon-poor environments-as the consequence of trade-offs between nutrient transport systems. While prototypical copiotrophs, like Vibrios, possess numerous phosphotransferase systems (PTS), prototypical oligotrophs, such as SAR11, lack PTS and rely on ATP-binding cassette (ABC) transporters, which use binding proteins. We develop models of both transport systems and use them in proteome allocation problems to predict the optimal nutrient uptake and metabolic strategy as a function of carbon availability. We derive a Michaelis-Menten approximation of ABC transport, analytically demonstrating how the half-saturation concentration is a function of binding protein abundance. We predict that oligotrophs can attain nanomolar half-saturation concentrations using binding proteins with only micromolar dissociation constants and while closely matching transport and metabolic capacities. However, our model predicts that this requires large periplasms and that the slow diffusion of the binding proteins limits uptake. Thus, binding proteins are critical for oligotrophic survival yet severely constrain growth rates. We propose that this trade-off fundamentally shaped the divergent evolution of oligotrophs and copiotrophs.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
