

Transcriptional co-activator regulates melanocyte differentiation and oncogenesis by integrating cAMP and MAPK/ERK pathways
- PMID: 34010639
- PMCID: PMC9678241
- DOI: 10.1016/j.celrep.2021.109136
Transcriptional co-activator regulates melanocyte differentiation and oncogenesis by integrating cAMP and MAPK/ERK pathways
Abstract
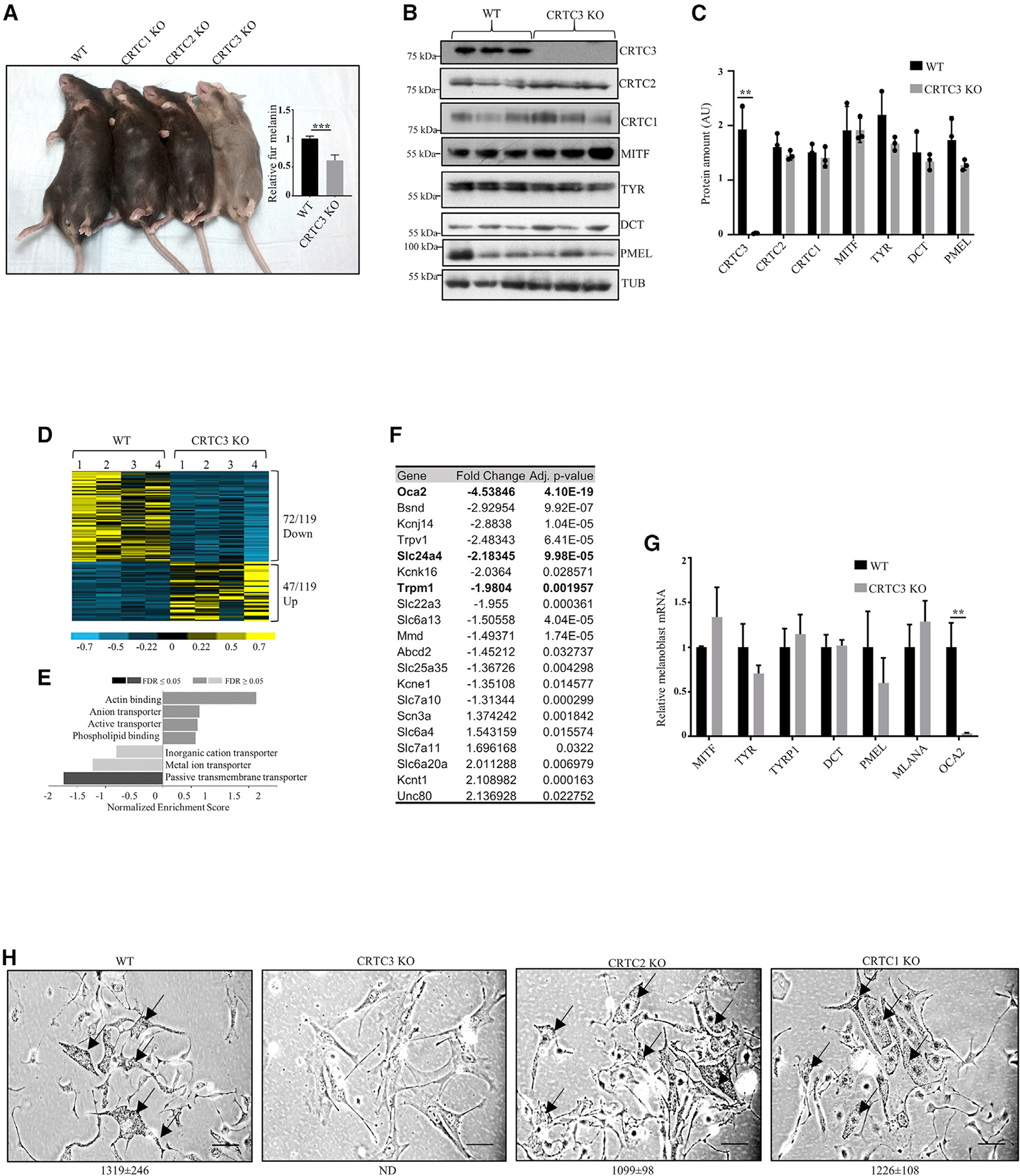
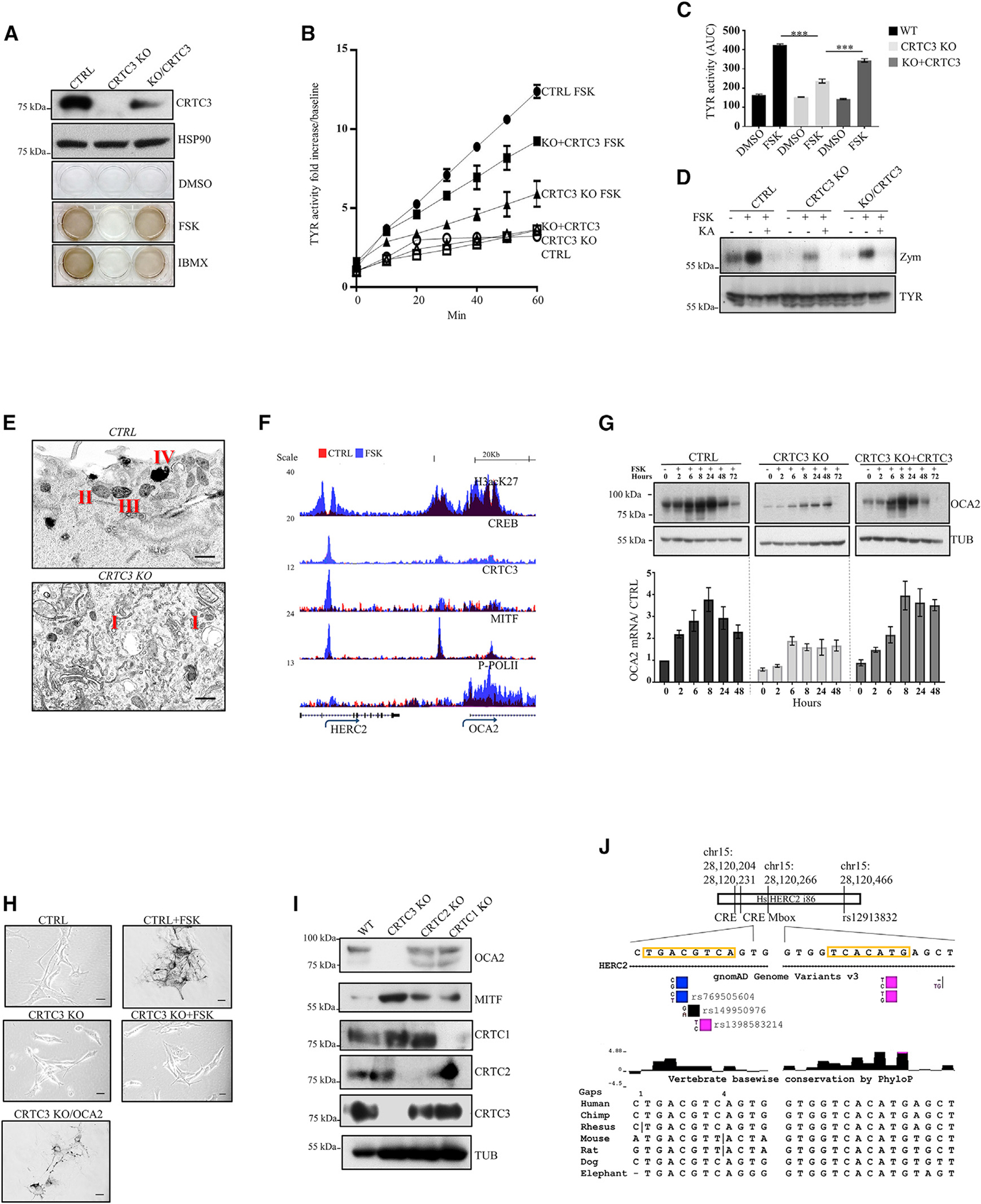
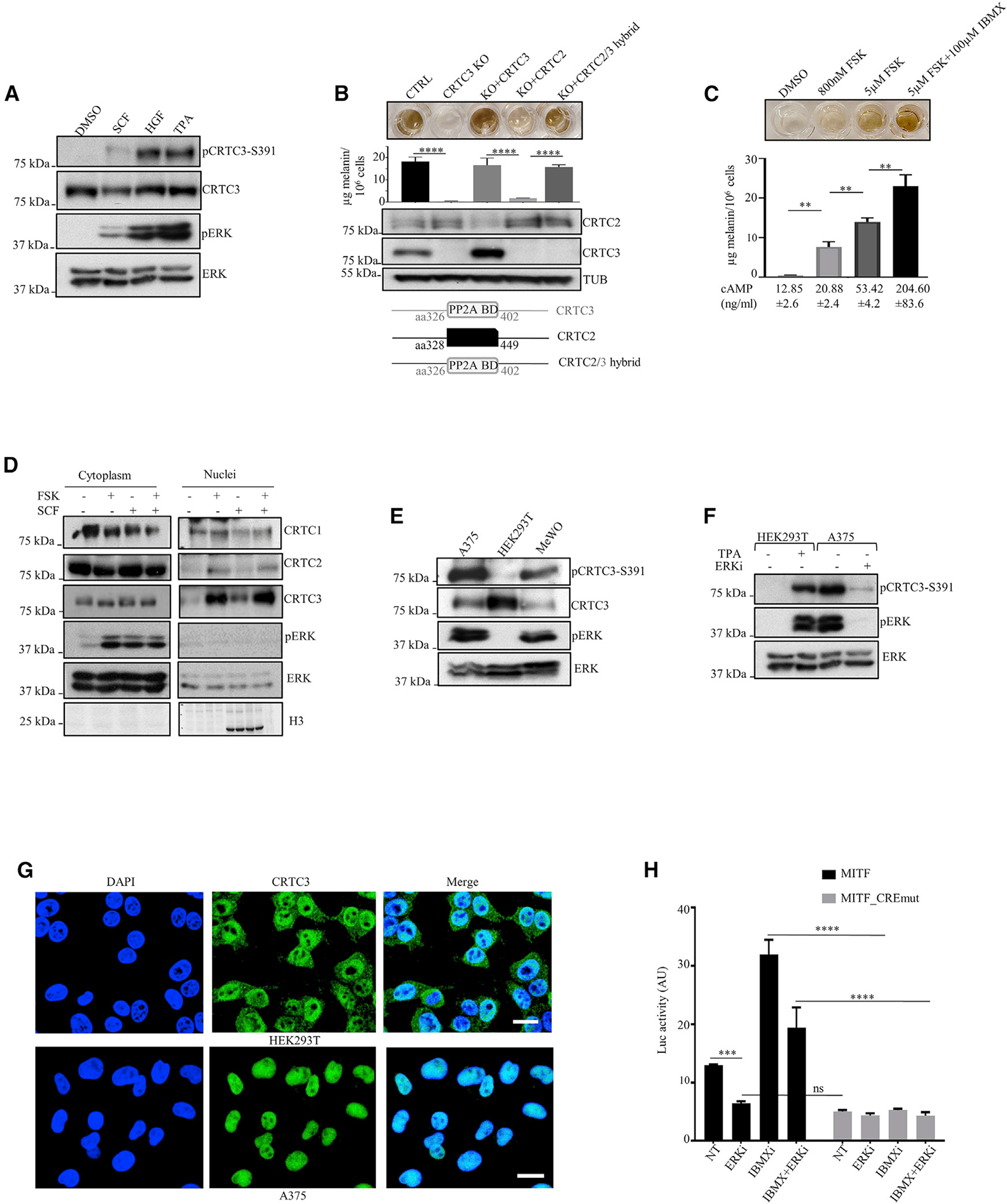
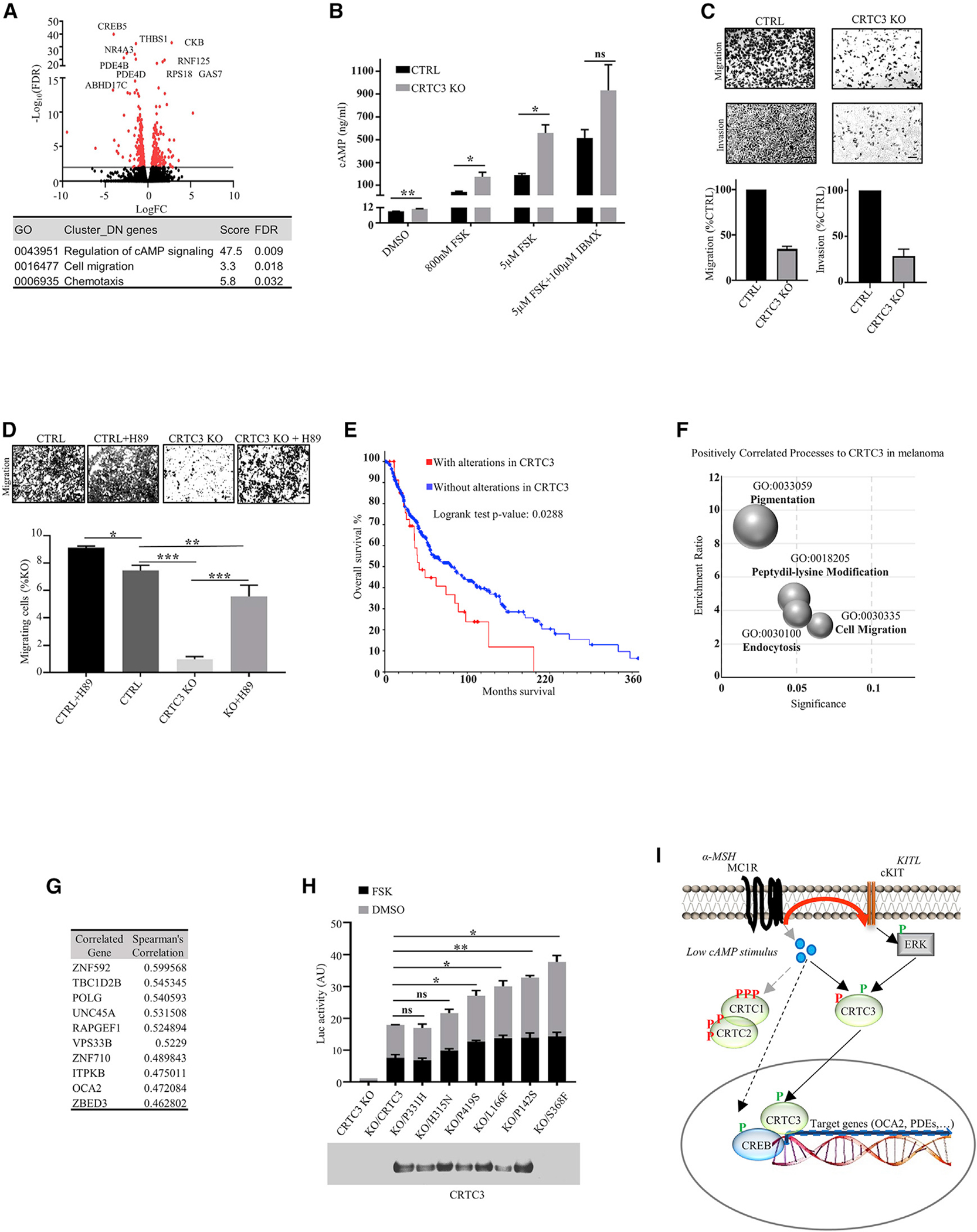
The cyclic AMP pathway promotes melanocyte differentiation by activating CREB and the cAMP-regulated transcription co-activators 1-3 (CRTC1-3). Differentiation is dysregulated in melanomas, although the contributions of CRTC proteins is unclear. We report a selective differentiation impairment in CRTC3 KO melanocytes and melanoma cells, due to downregulation of oculo-cutaneous albinism II (OCA2) and block of melanosome maturation. CRTC3 stimulates OCA2 expression by binding to CREB on a conserved enhancer, a regulatory site for pigmentation and melanoma risk. CRTC3 is uniquely activated by ERK1/2-mediated phosphorylation at Ser391 and by low levels of cAMP. Phosphorylation at Ser391 is constitutively elevated in human melanoma cells with hyperactivated ERK1/2 signaling; knockout of CRTC3 in this setting impairs anchorage-independent growth, migration, and invasiveness, whereas CRTC3 overexpression supports cell survival in response to the mitogen-activated protein kinase (MAPK) inhibitor vemurafenib. As melanomas expressing gain-of-function mutations in CRTC3 are associated with reduced survival, our results suggest that CRTC3 inhibition may provide therapeutic benefit in this setting.
Keywords: CREB; CRTC3; ERK; OCA2; cAMP; melanoma; melanosome; phospho-diesterase; pigmentation.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Bang S, Won KH, Moon H-R, Yoo H, Hong A, Song Y, and Chang SE (2017). Novel regulation of melanogenesis by adiponectin via the AMPK/CRTC pathway. Pigment Cell Melanoma Res. 30, 553–557. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
