

Cortex-dependent corrections as the tongue reaches for and misses targets
- PMID: 34012117
- PMCID: PMC8299742
- DOI: 10.1038/s41586-021-03561-9
Cortex-dependent corrections as the tongue reaches for and misses targets
Abstract
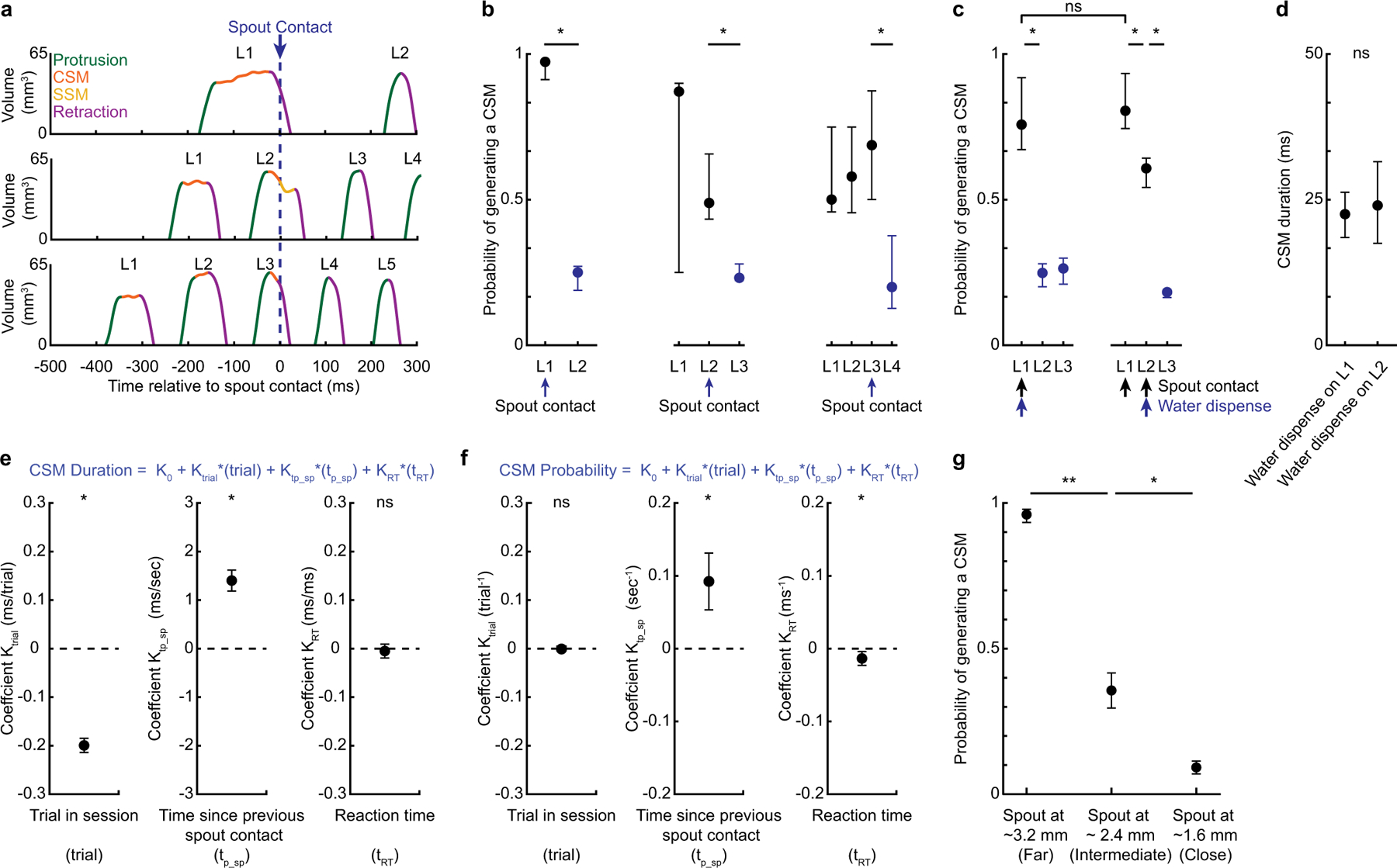
Precise tongue control is necessary for drinking, eating and vocalizing1-3. However, because tongue movements are fast and difficult to resolve, neural control of lingual kinematics remains poorly understood. Here we combine kilohertz-frame-rate imaging and a deep-learning-based neural network to resolve 3D tongue kinematics in mice drinking from a water spout. Successful licks required corrective submovements that-similar to online corrections during primate reaches4-11-occurred after the tongue missed unseen, distant or displaced targets. Photoinhibition of anterolateral motor cortex impaired corrections, which resulted in hypometric licks that missed the spout. Neural activity in anterolateral motor cortex reflected upcoming, ongoing and past corrective submovements, as well as errors in predicted spout contact. Although less than a tenth of a second in duration, a single mouse lick exhibits the hallmarks of online motor control associated with a primate reach, including cortex-dependent corrections after misses.
Conflict of interest statement
Figures













References
-
- Kier WM & Smith KK Tongues, tentacles and trunks: the biomechanics of movement in muscular-hydrostats. Zool. J. Linn. Soc 83, 307–324 (1985).
-
- Meyer DE, Abrams RA, Kornblum S, Wright CE & Smith JE Optimality in human motor performance: ideal control of rapid aimed movements. Psychol. Rev. 95, 340–370 (1988). - PubMed
-
- Spijkers WA & Lochner P Partial visual feedback and spatial end-point accuracy of discrete aiming movements. J. Mot. Behav 26, 283–295 (1994). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
