

Treatment with hypoxia-mimetics protects cultured rat Schwann cells against oxidative stress-induced cell death
- PMID: 34019306
- PMCID: PMC11848739
- DOI: 10.1002/glia.24019
Treatment with hypoxia-mimetics protects cultured rat Schwann cells against oxidative stress-induced cell death
Abstract
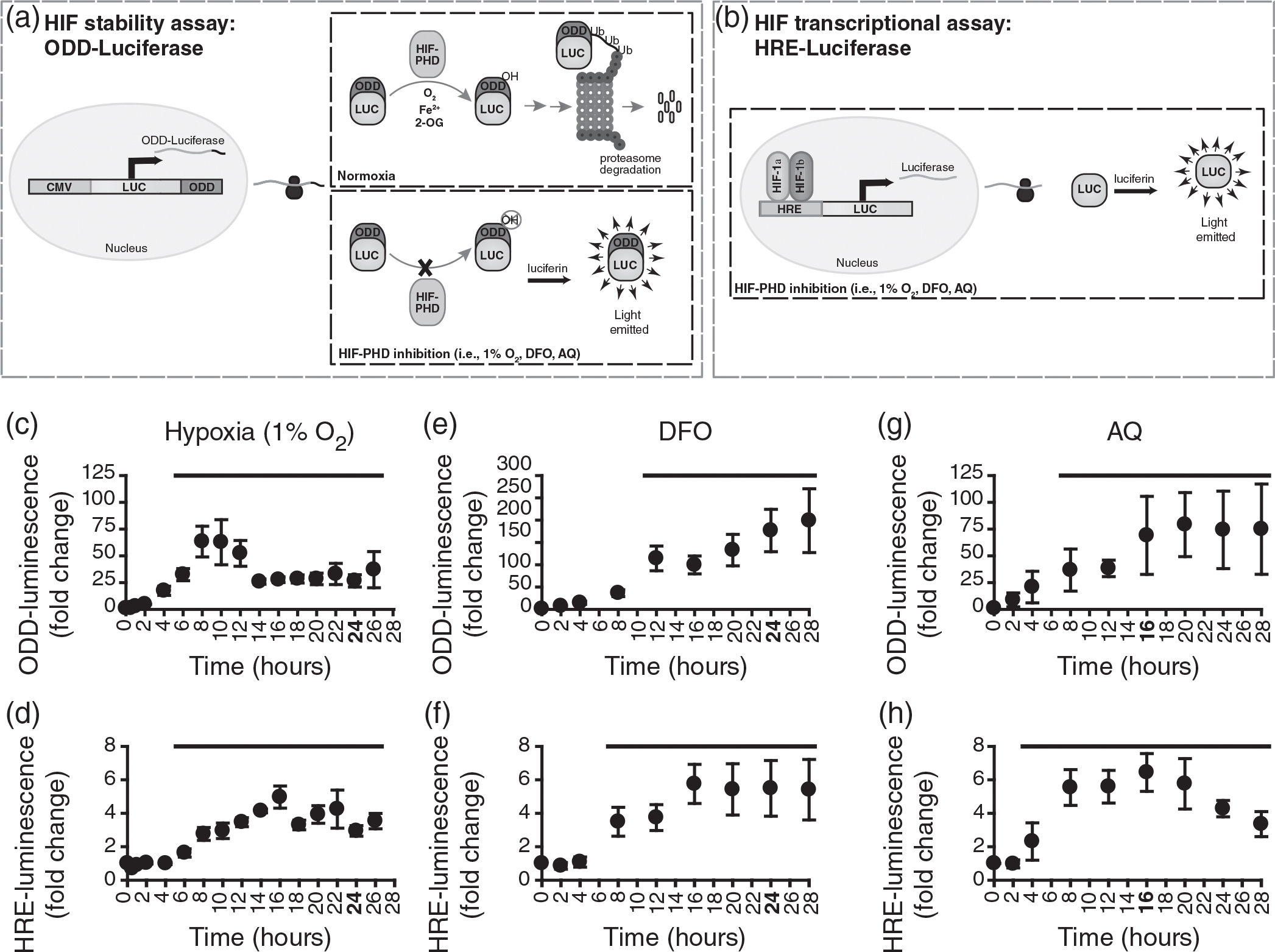
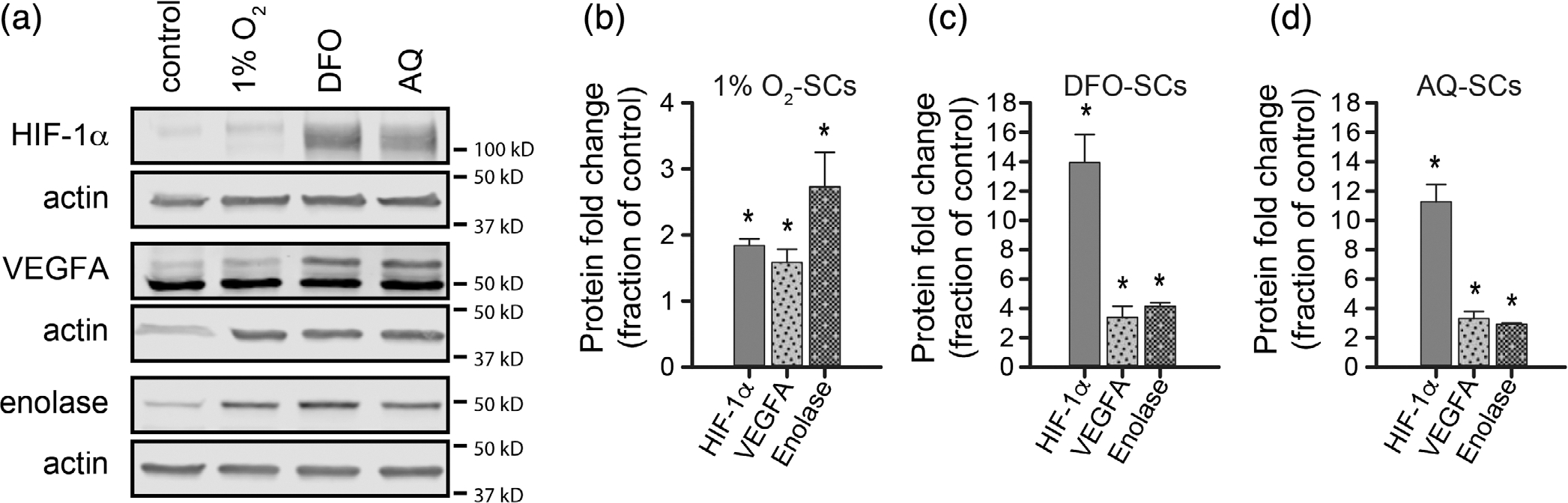
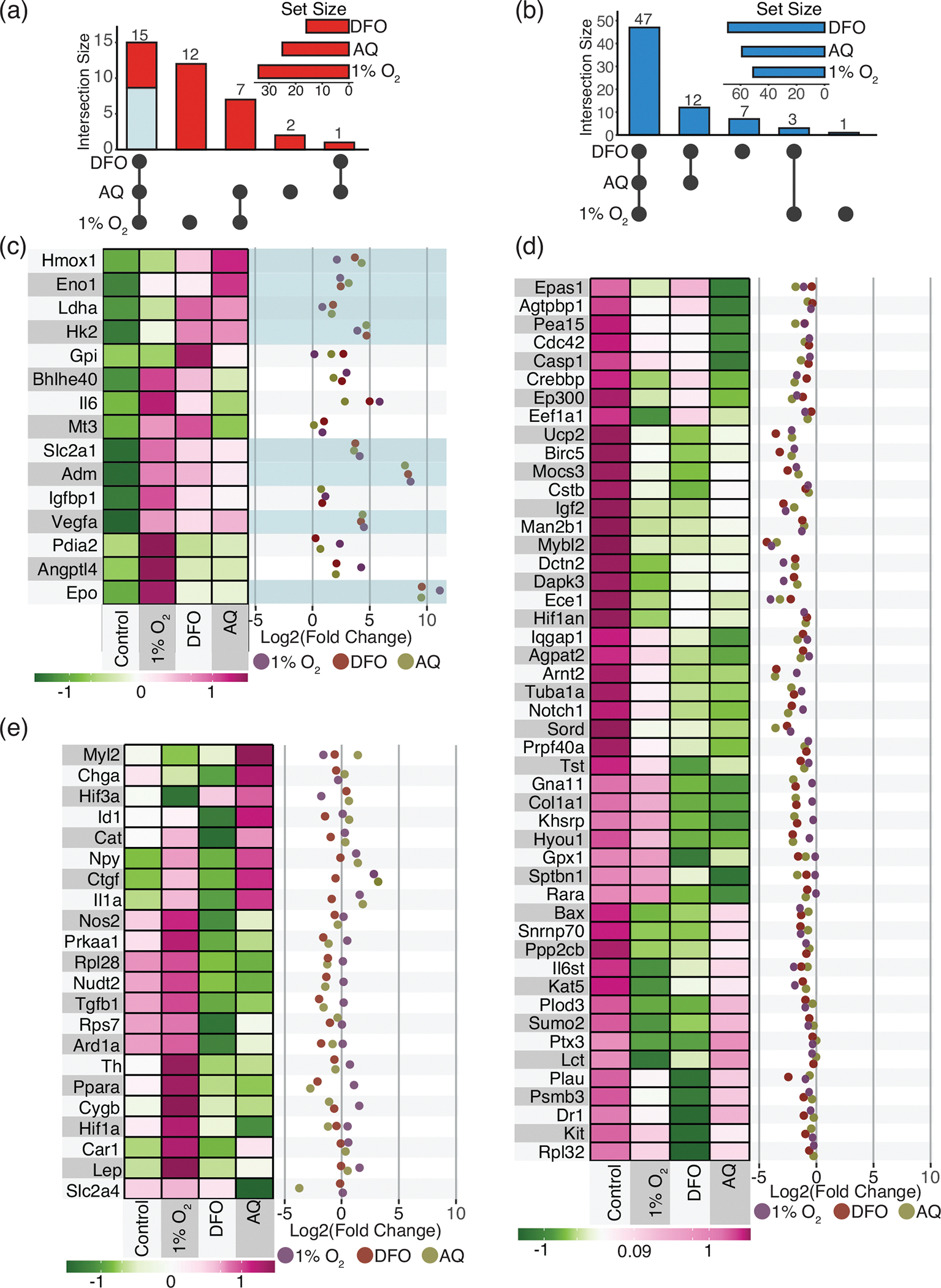
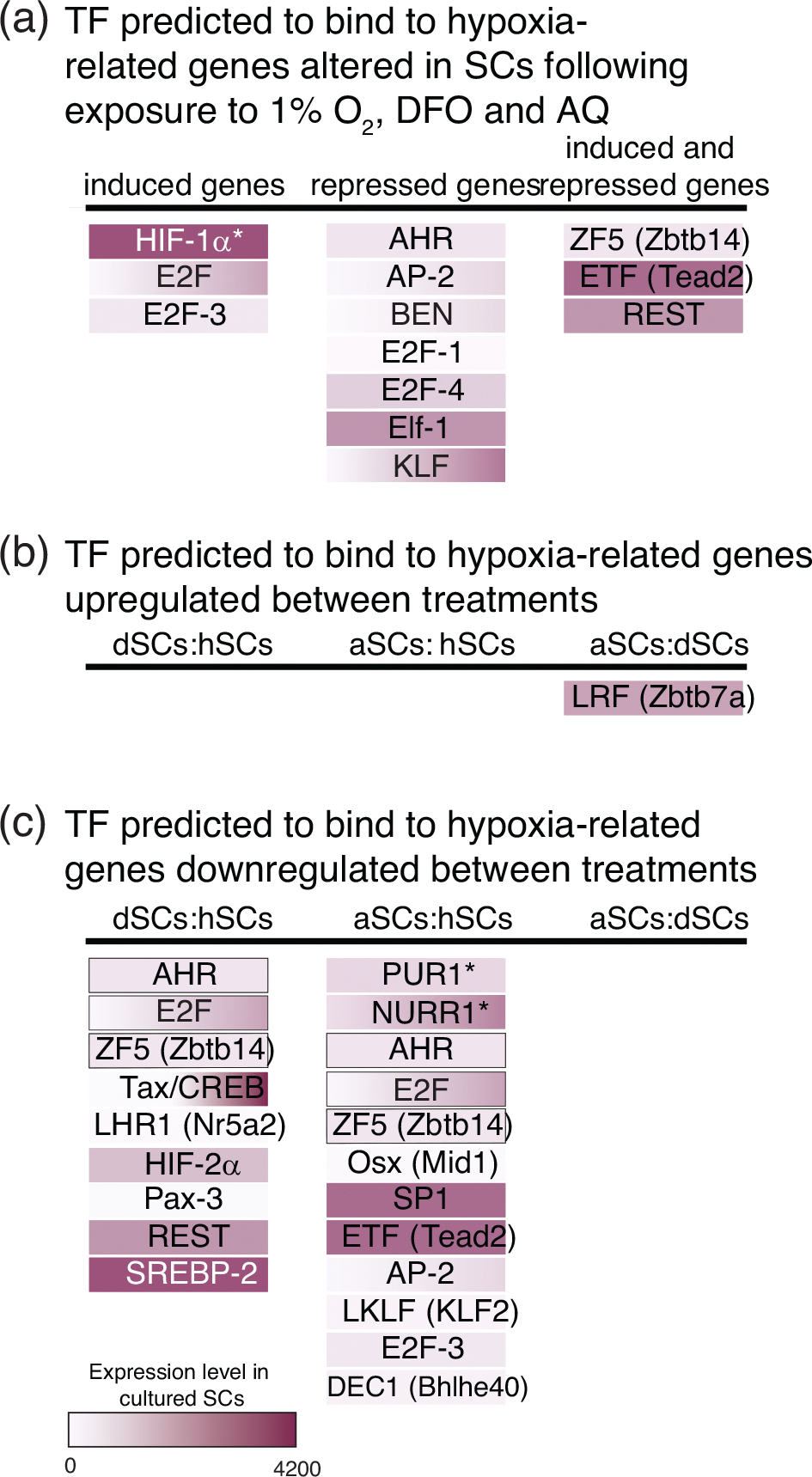
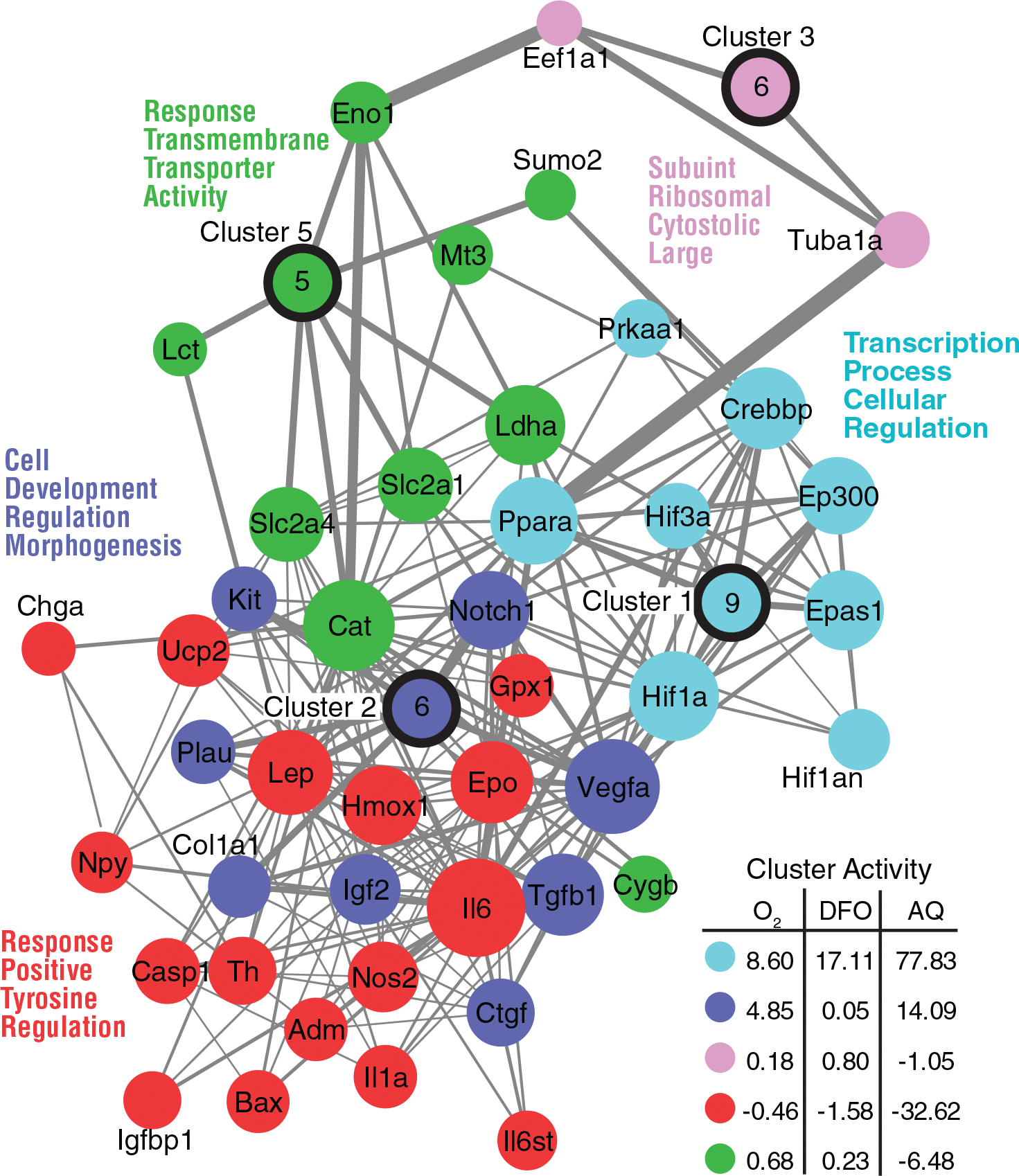
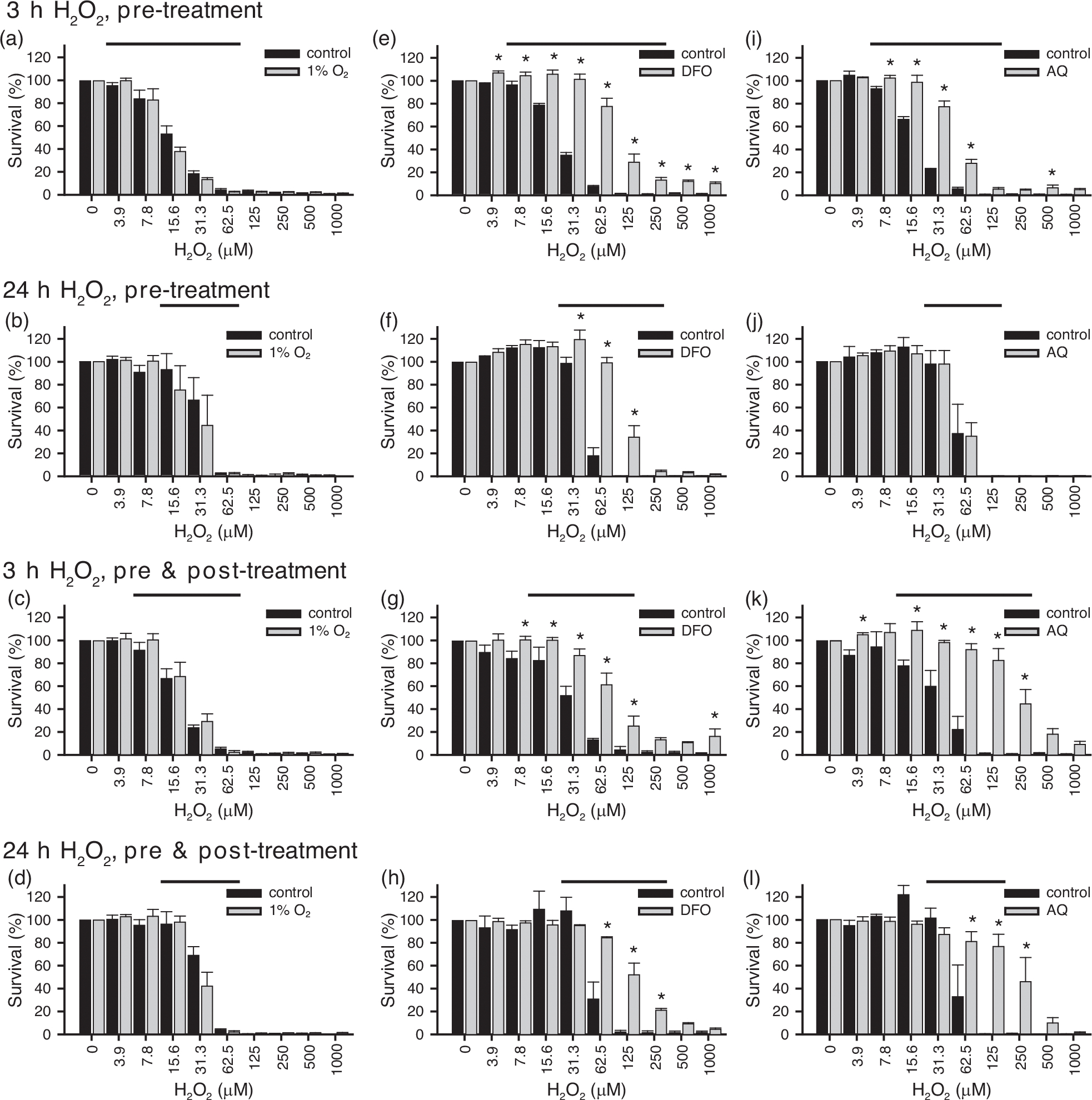
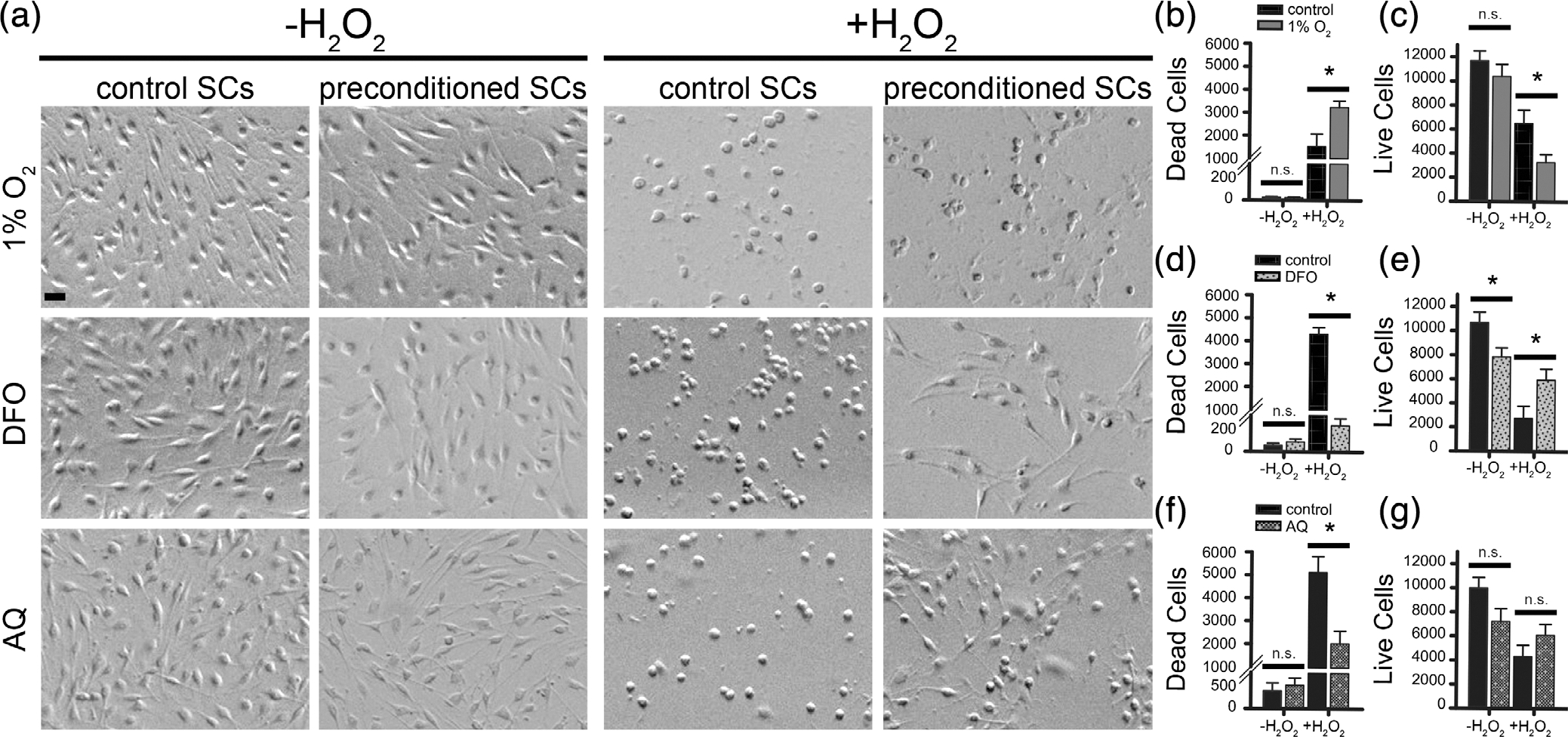
Schwann cell (SC) grafts promote axon regeneration in the injured spinal cord, but transplant efficacy is diminished by a high death rate in the first 2-3 days postimplantation. Both hypoxic preconditioning and pharmacological induction of the cellular hypoxic response can drive cellular adaptations and improve transplant survival in a number of disease/injury models. Hypoxia-inducible factor 1 alpha (HIF-1α), a regulator of the cellular response to hypoxia, is implicated in preconditioning-associated protection. HIF-1α cellular levels are regulated by the HIF-prolyl hydroxylases (HIF-PHDs). Pharmacological inhibition of the HIF-PHDs mimics hypoxic preconditioning and provides a method to induce adaptive hypoxic responses without direct exposure to hypoxia. In this study, we show that hypoxia-mimetics, deferoxamine (DFO) and adaptaquin (AQ), enhance HIF-1α stability and HIF-1α target gene expression. Expression profiling of hypoxia-related genes demonstrates that HIF-dependent and HIF-independent expression changes occur. Analyses of transcription factor binding sites identify several candidate transcriptional co-regulators that vary in SCs along with HIF-1α. Using an in vitro model system, we show that hypoxia-mimetics are potent blockers of oxidative stress-induced death in SCs. In contrast, traditional hypoxic preconditioning was not protective. The robust protection induced by pharmacological preconditioning, particularly with DFO, indicates that pharmacological induction of hypoxic adaptations could be useful for promoting transplanted SC survival. These agents may also be more broadly useful for protecting SCs, as oxidative stress is a major pathway that drives cellular damage in the context of neurological injury and disease, including demyelinating diseases and peripheral neuropathies.
Keywords: H2O2; adaptaquin; cell death; cell survival; deferoxamine; hypoxia adaptations; hypoxia inducible factor (HIF); preconditioning; prolyl hydroxylase inhibition; reporter assay.
© 2021 Wiley Periodicals LLC.
Conflict of interest statement
CONFLICT OF INTEREST
The authors declare no conflicts of interest.
Figures
Similar articles
-
Temporary induction of hypoxic adaptations by preconditioning fails to enhance Schwann cell transplant survival after spinal cord injury.Glia. 2023 Mar;71(3):648-666. doi: 10.1002/glia.24302. Epub 2022 Dec 24. Glia. 2023. PMID: 36565279 Free PMC article.
-
Hypoxia-Inducible Factor 1α (HIF-1α) Counteracts the Acute Death of Cells Transplanted into the Injured Spinal Cord.eNeuro. 2020 May 11;7(3):ENEURO.0092-19.2019. doi: 10.1523/ENEURO.0092-19.2019. Print 2020 May/Jun. eNeuro. 2020. PMID: 31488552 Free PMC article.
-
Hypoxic preconditioning produces differential expression of hypoxia-inducible factor-1alpha (HIF-1alpha) and its regulatory enzyme HIF prolyl hydroxylase 2 in neonatal rat brain.Neurosci Lett. 2006 Aug 14;404(1-2):72-7. doi: 10.1016/j.neulet.2006.05.049. Epub 2006 Jul 7. Neurosci Lett. 2006. PMID: 16828226
-
Hypoxia Mimetic Agents for Ischemic Stroke.Front Cell Dev Biol. 2019 Jan 8;6:175. doi: 10.3389/fcell.2018.00175. eCollection 2018. Front Cell Dev Biol. 2019. PMID: 30671433 Free PMC article. Review.
-
Extracellular Vesicles as Transmitters of Hypoxia Tolerance in Solid Cancers.Cancers (Basel). 2019 Jan 29;11(2):154. doi: 10.3390/cancers11020154. Cancers (Basel). 2019. PMID: 30699970 Free PMC article. Review.
Cited by
-
Enhanced sciatic nerve regeneration by relieving iron-overloading and organelle stress with the nanofibrous P(MMD-co-LA)/DFO conduits.Mater Today Bio. 2022 Aug 6;16:100387. doi: 10.1016/j.mtbio.2022.100387. eCollection 2022 Dec. Mater Today Bio. 2022. PMID: 36042854 Free PMC article.
-
Temporary induction of hypoxic adaptations by preconditioning fails to enhance Schwann cell transplant survival after spinal cord injury.Glia. 2023 Mar;71(3):648-666. doi: 10.1002/glia.24302. Epub 2022 Dec 24. Glia. 2023. PMID: 36565279 Free PMC article.
-
Enhancing intraneural revascularization following peripheral nerve injury through hypoxic Schwann-cell-derived exosomes: an insight into endothelial glycolysis.J Nanobiotechnology. 2024 May 24;22(1):283. doi: 10.1186/s12951-024-02536-y. J Nanobiotechnology. 2024. PMID: 38789980 Free PMC article.
-
Efficacy of Deferoxamine Mesylate in Serum and Serum-Free Media: Adult Ventral Root Schwann Cell Survival Following Hydrogen Peroxide-Induced Cell Death.Cells. 2025 Mar 20;14(6):461. doi: 10.3390/cells14060461. Cells. 2025. PMID: 40136710 Free PMC article.
-
Mesenchymal Stem Cell-Derived Extracellular Vesicles: A Potential Therapy for Diabetes Mellitus and Diabetic Complications.Pharmaceutics. 2022 Oct 17;14(10):2208. doi: 10.3390/pharmaceutics14102208. Pharmaceutics. 2022. PMID: 36297643 Free PMC article. Review.
References
-
- Aime P, Karuppagounder SS, Rao A, Chen Y, Burke RE, Ratan RR, & Greene LA (2020). The drug adaptaquin blocks ATF4/CHOP-dependent pro-death Trib3 induction and protects in cellular and mouse models of Parkinson’s disease. Neurobiology of Disease, 136, 104725. 10.1016/j.nbd.2019.104725 - DOI - PMC - PubMed
-
- Anderson KD, Guest JD, Dietrich WD, Bartlett Bunge M, Curiel R, Dididze M, Green BA, Khan A, Pearse DD, Saraf-Lavi E, Widerstrom-Noga E, Wood P, & Levi AD (2017). Safety of autologous human Schwann cell transplantation in subacute thoracic spinal cord injury. Journal of Neurotrauma, 34(21), 2950–2963. 10.1089/neu.2016.4895 - DOI - PubMed
-
- Aoki H, & Kunisada T (2016). Rest as a new transcription factor to control neural crest development. Single Cell Biology, 5(2). 10.4172/2168-9431.1000144 - DOI
-
- Appelhoff RJ, Tian YM, Raval RR, Turley H, Harris AL, Pugh CW, Ratcliffe PJ, & Gleadle JM (2004). Differential function of the prolyl hydroxylases PHD1, PHD2, and PHD3 in the regulation of hypoxia-inducible factor. The Journal of Biological Chemistry, 279(37), 38458–38465. 10.1074/jbc.M406026200 - DOI - PubMed
-
- Arany Z, Huang LE, Eckner R, Bhattacharya S, Jiang C, Goldberg MA, Bunn HF, & Livingston DM (1996). An essential role for p300/CBP in the cellular response to hypoxia. Proceedings of the National Academy of Sciences of the United States of America, 93 (23), 12969–12973. 10.1073/pnas.93.23.12969 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
