

Increased tissue stiffness triggers contractile dysfunction and telomere shortening in dystrophic cardiomyocytes
- PMID: 34019816
- PMCID: PMC8452491
- DOI: 10.1016/j.stemcr.2021.04.018
Increased tissue stiffness triggers contractile dysfunction and telomere shortening in dystrophic cardiomyocytes
Abstract
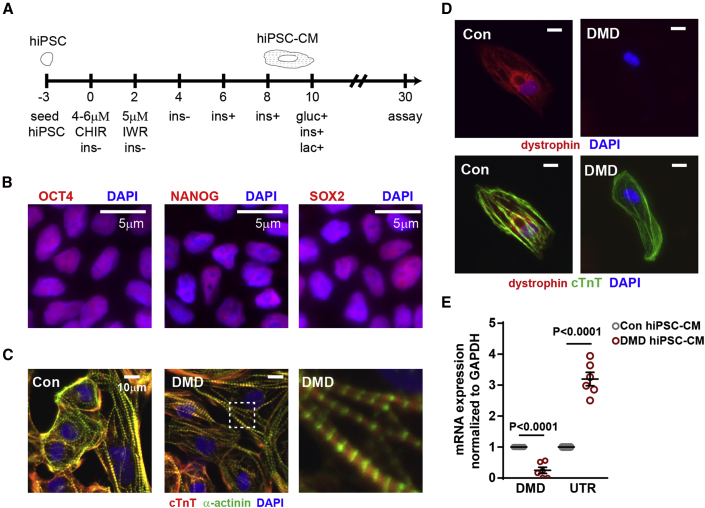
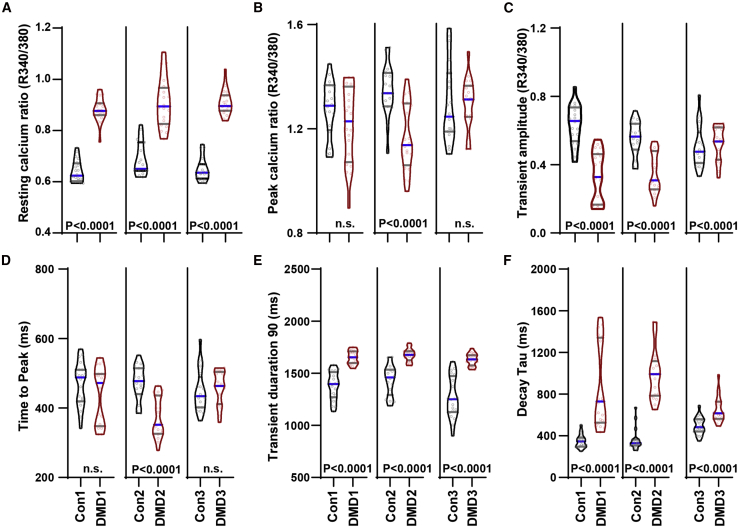
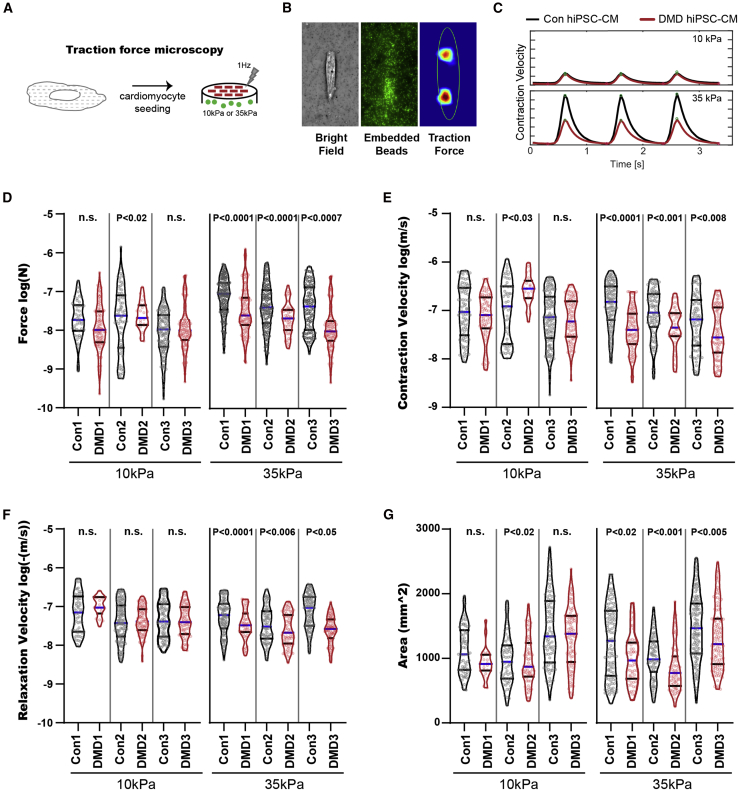
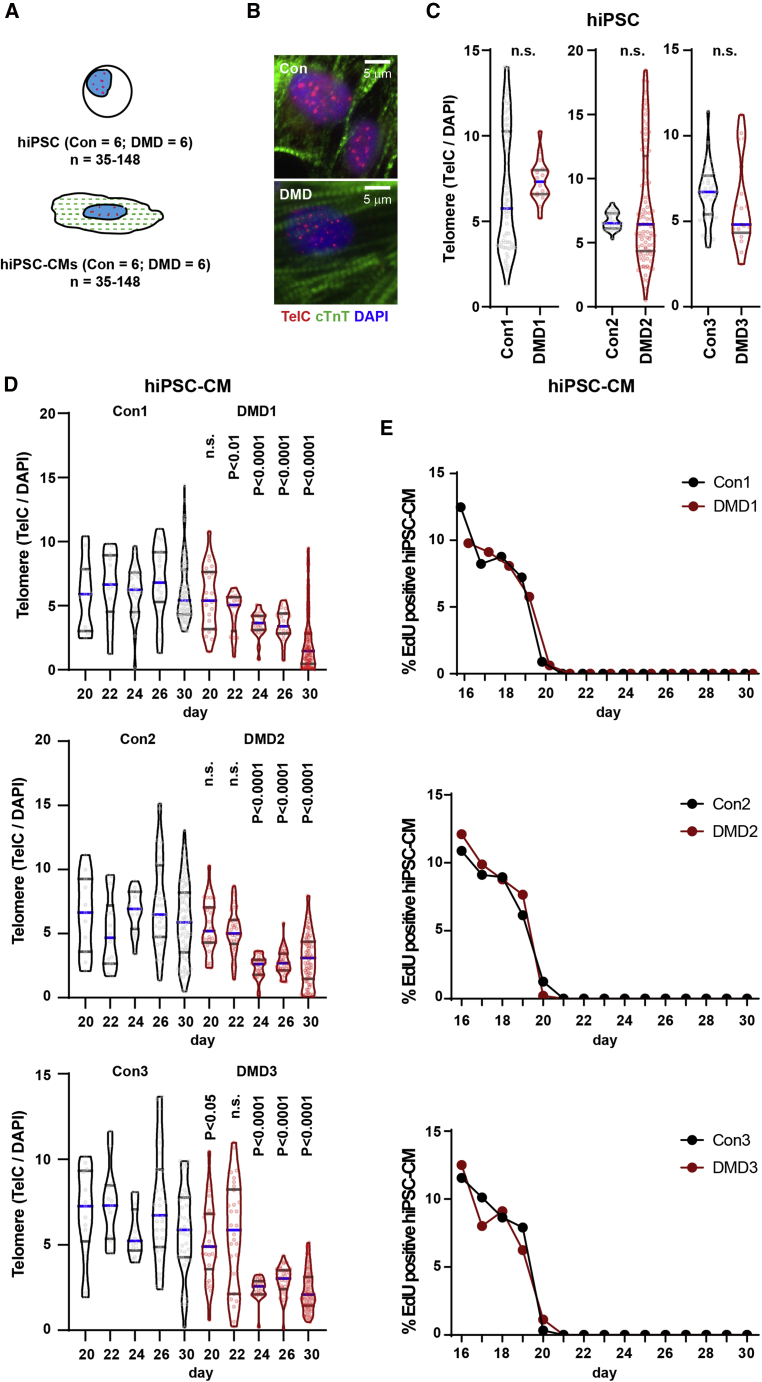
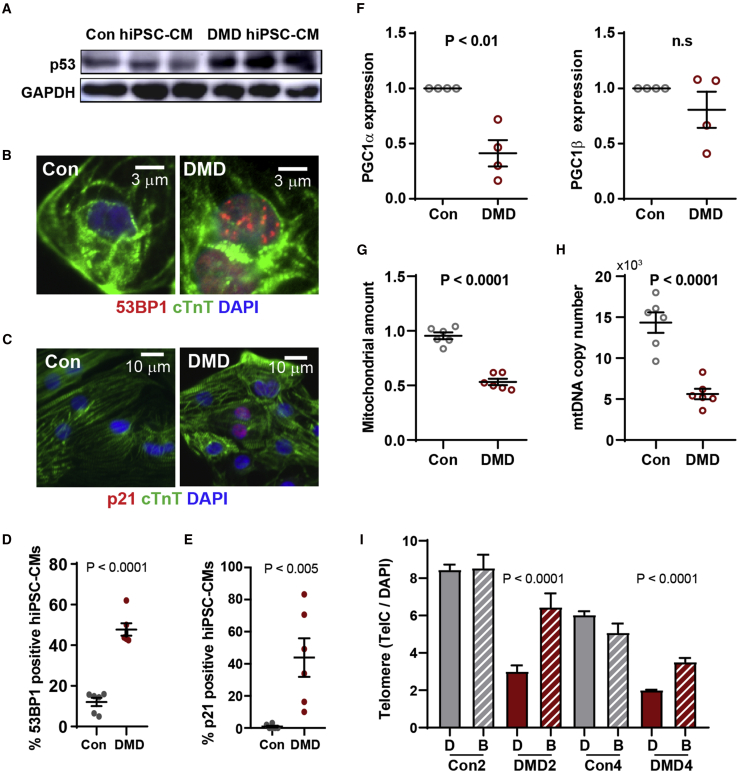
Duchenne muscular dystrophy (DMD) is a rare X-linked recessive disease that is associated with severe progressive muscle degeneration culminating in death due to cardiorespiratory failure. We previously observed an unexpected proliferation-independent telomere shortening in cardiomyocytes of a DMD mouse model. Here, we provide mechanistic insights using human induced pluripotent stem cell-derived cardiomyocytes (hiPSC-CMs). Using traction force microscopy, we show that DMD hiPSC-CMs exhibit deficits in force generation on fibrotic-like bioengineered hydrogels, aberrant calcium handling, and increased reactive oxygen species levels. Furthermore, we observed a progressive post-mitotic telomere shortening in DMD hiPSC-CMs coincident with downregulation of shelterin complex, telomere capping proteins, and activation of the p53 DNA damage response. This telomere shortening is blocked by blebbistatin, which inhibits contraction in DMD cardiomyocytes. Our studies underscore the role of fibrotic stiffening in the etiology of DMD cardiomyopathy. In addition, our data indicate that telomere shortening is progressive, contraction dependent, and mechanosensitive, and suggest points of therapeutic intervention.
Keywords: DMD; dilated cardiomyopathy; fibrosis; hiPSC-CM; telomere.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Aguado J., Sola-Carvajal A., Cancila V., Revêchon G., Ong P.F., Jones-Weinert C.W., Wallén Arzt E., Lattanzi G., Dreesen O., Tripodo C., et al. Inhibition of DNA damage response at telomeres improves the detrimental phenotypes of Hutchinson-Gilford progeria syndrome. Nat. Commun. 2019;10:4990–5011. doi: 10.1038/s41467-019-13018-3. - DOI - PMC - PubMed
-
- Amoasii L., Hildyard J.C.W., Li H., Sanchez-Ortiz E., Mireault A., Caballero D., Harron R., Stathopoulou T.-R., Massey C., Shelton J.M., et al. Gene editing restores dystrophin expression in a canine model of Duchenne muscular dystrophy. Science. 2018;8:eaau1549. doi: 10.1126/science.aau1549. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
