

Adhesion-mediated mechanosignaling forces mitohormesis
- PMID: 34019840
- PMCID: PMC8266765
- DOI: 10.1016/j.cmet.2021.04.017
Adhesion-mediated mechanosignaling forces mitohormesis
Abstract
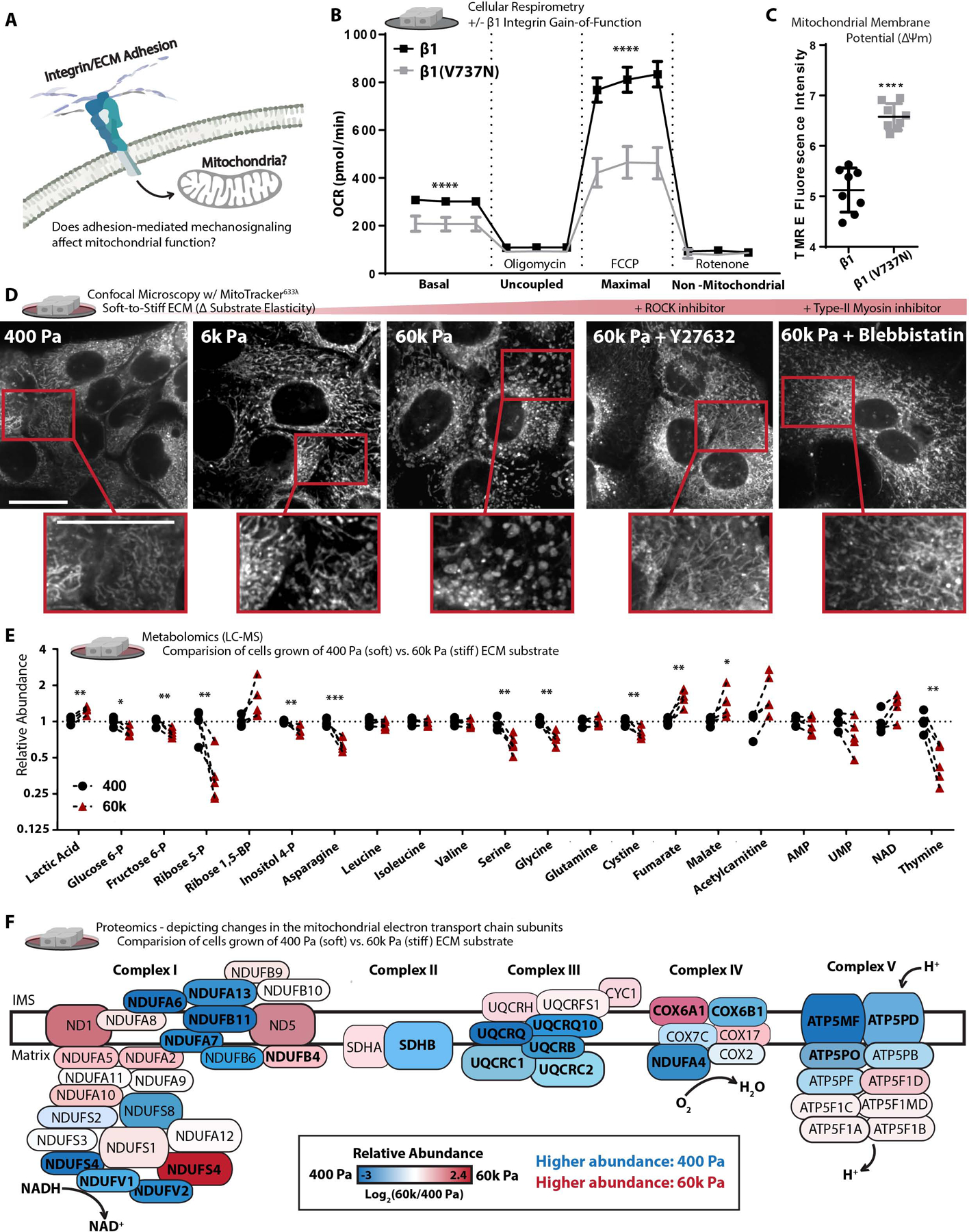
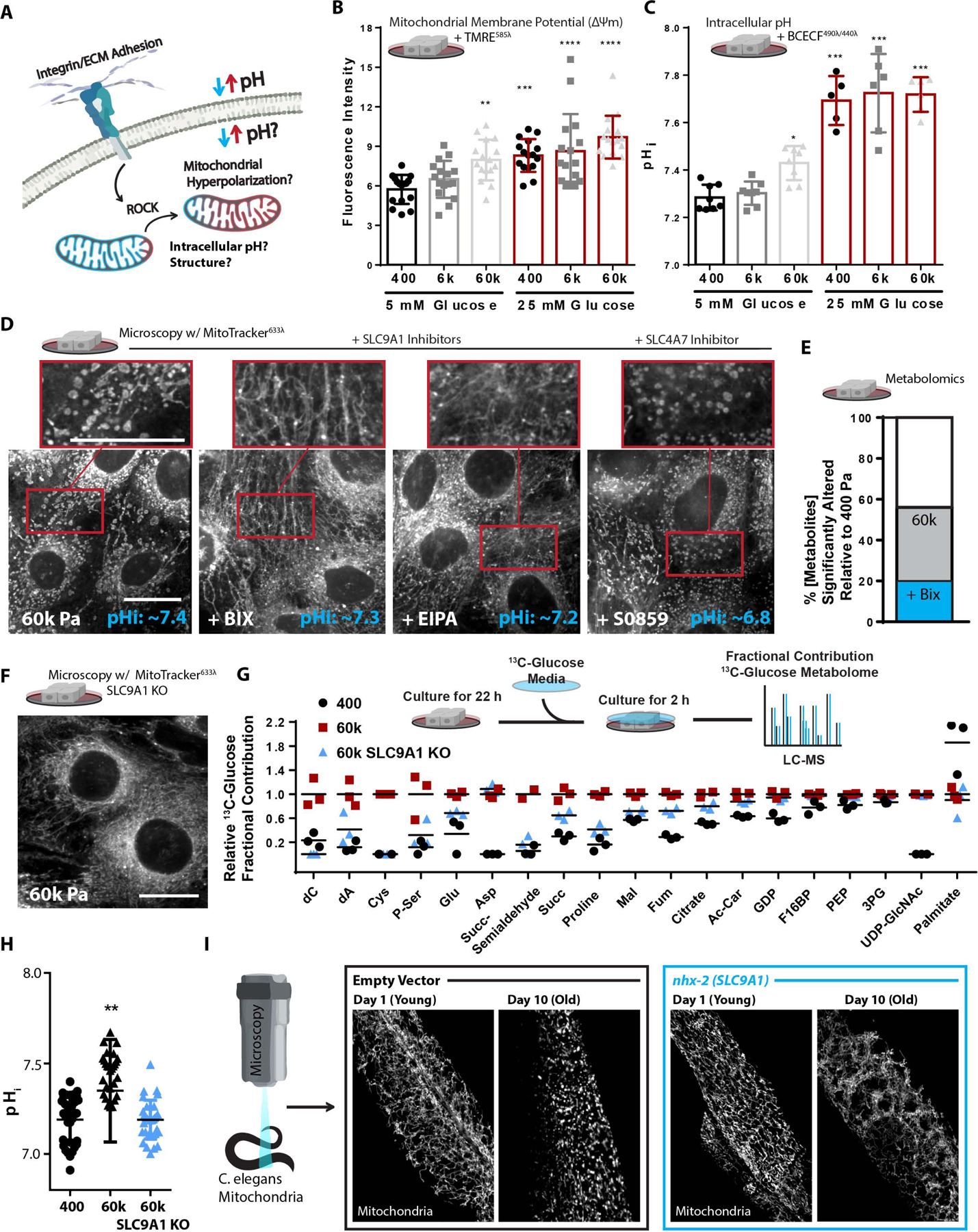
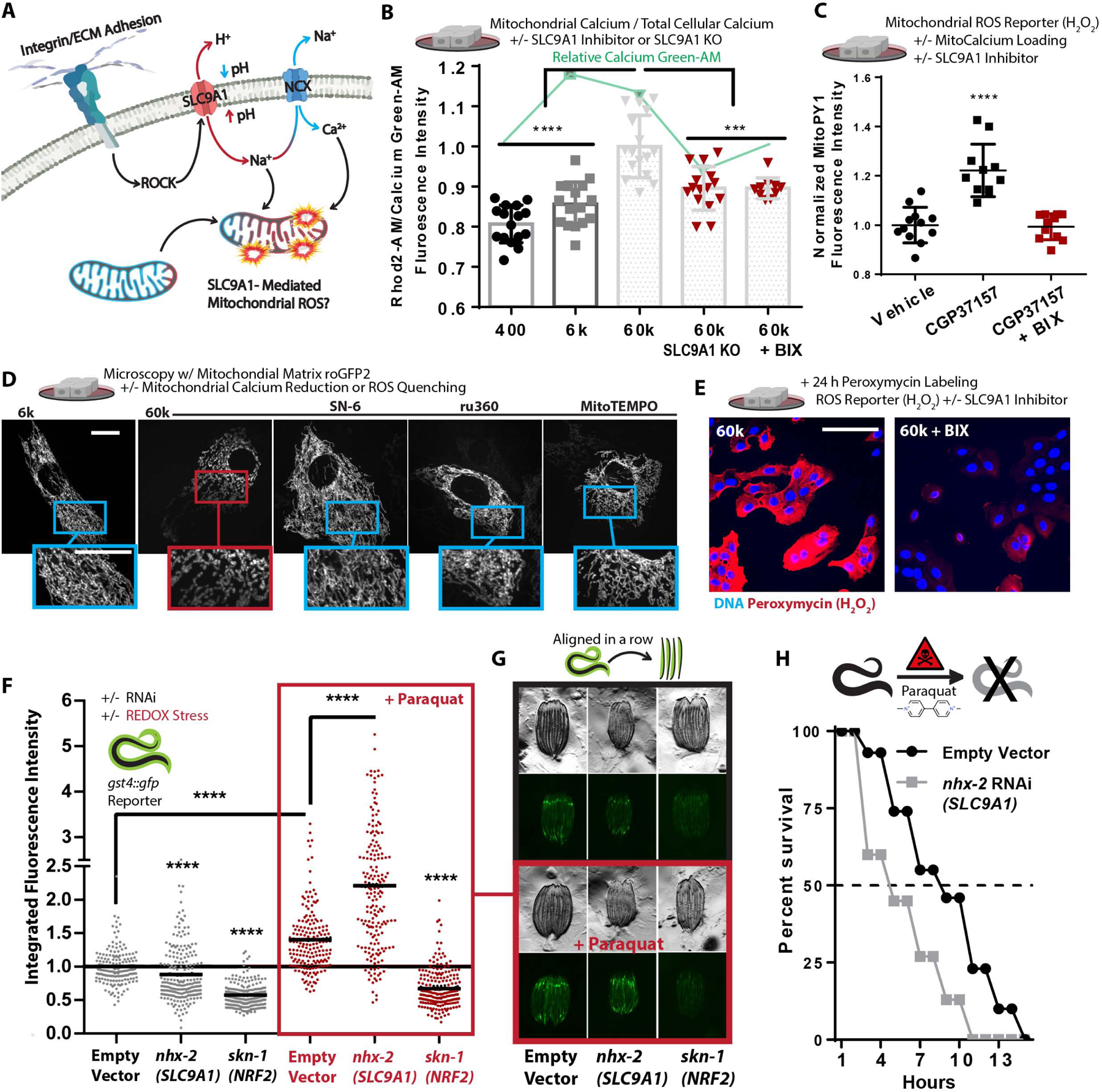
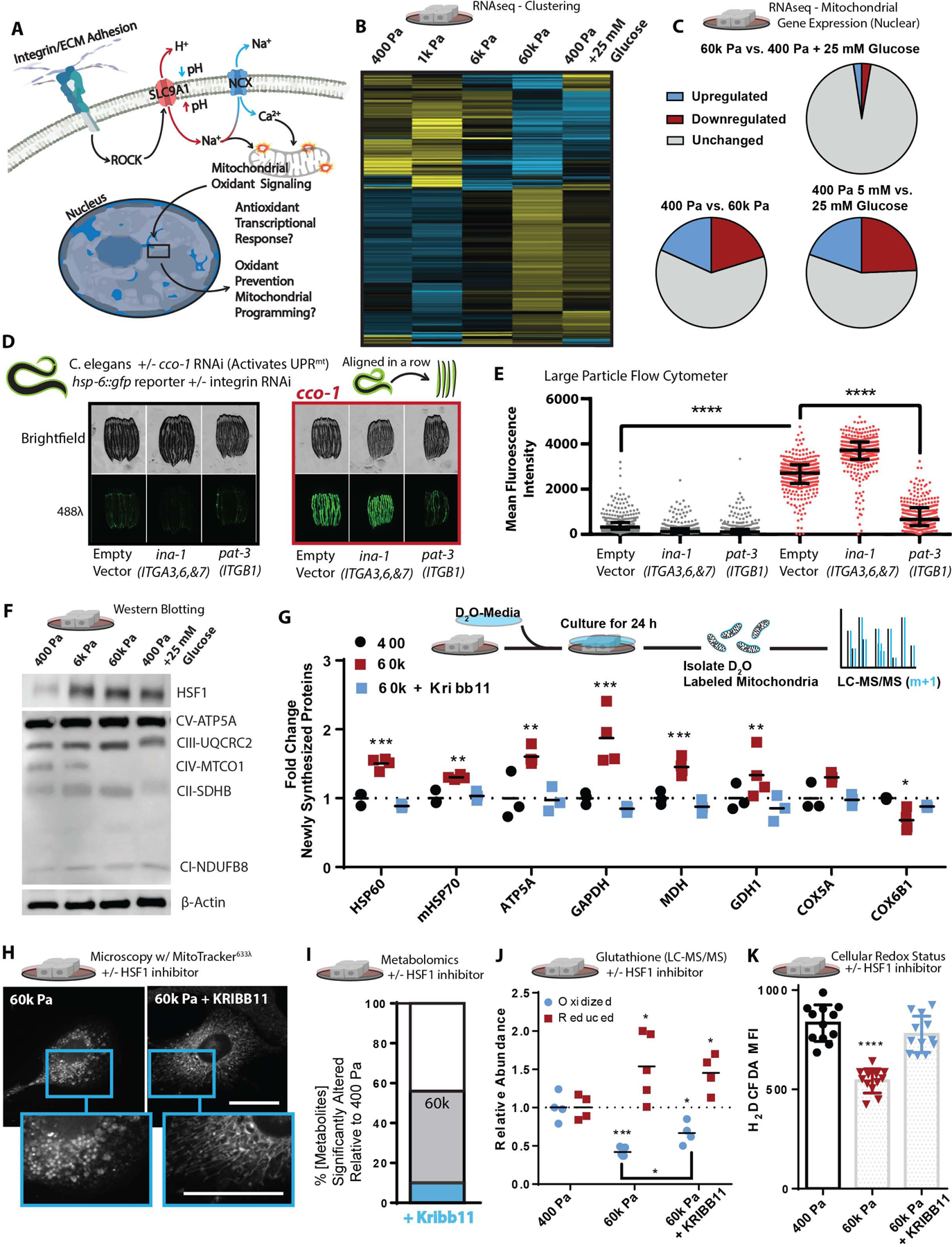
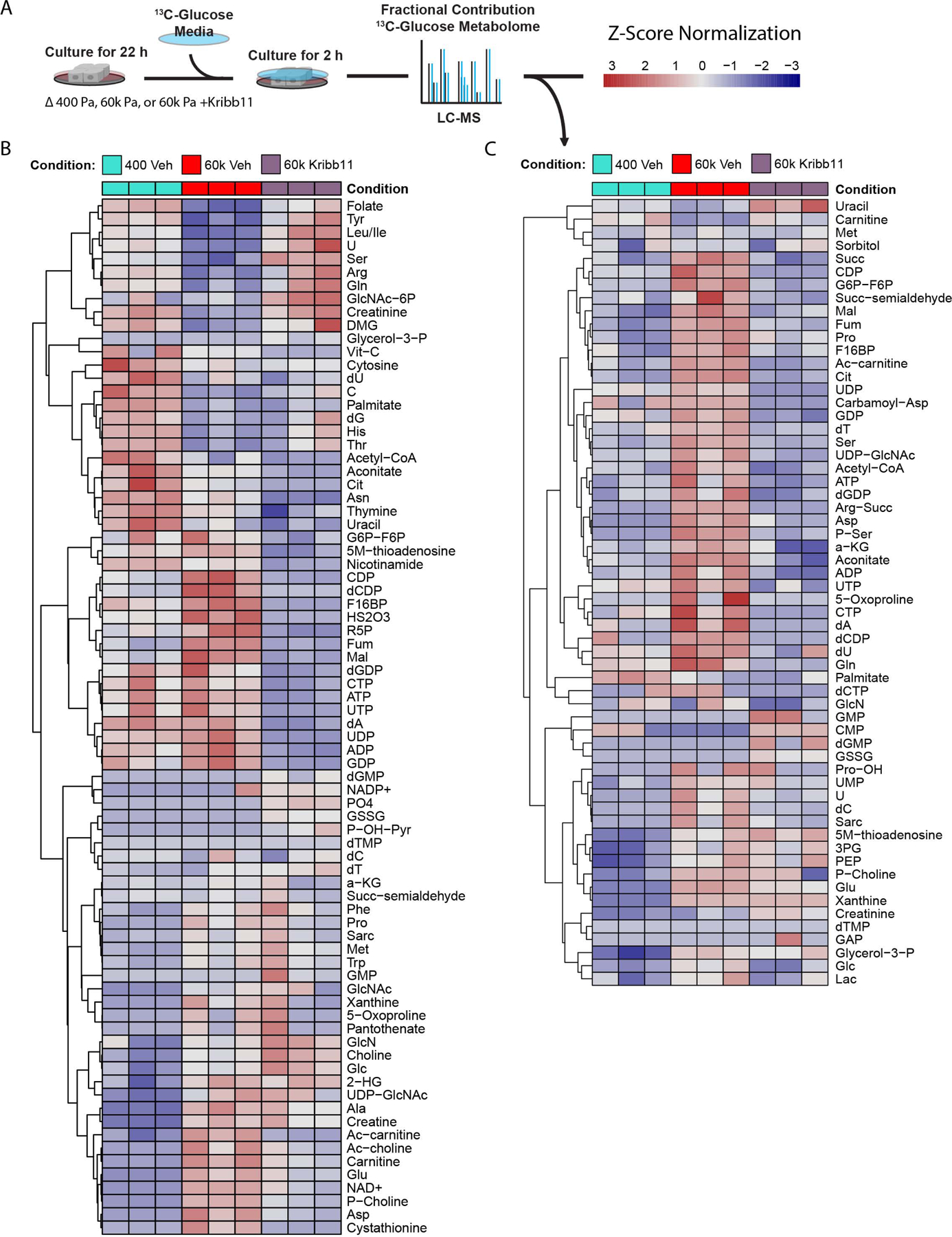
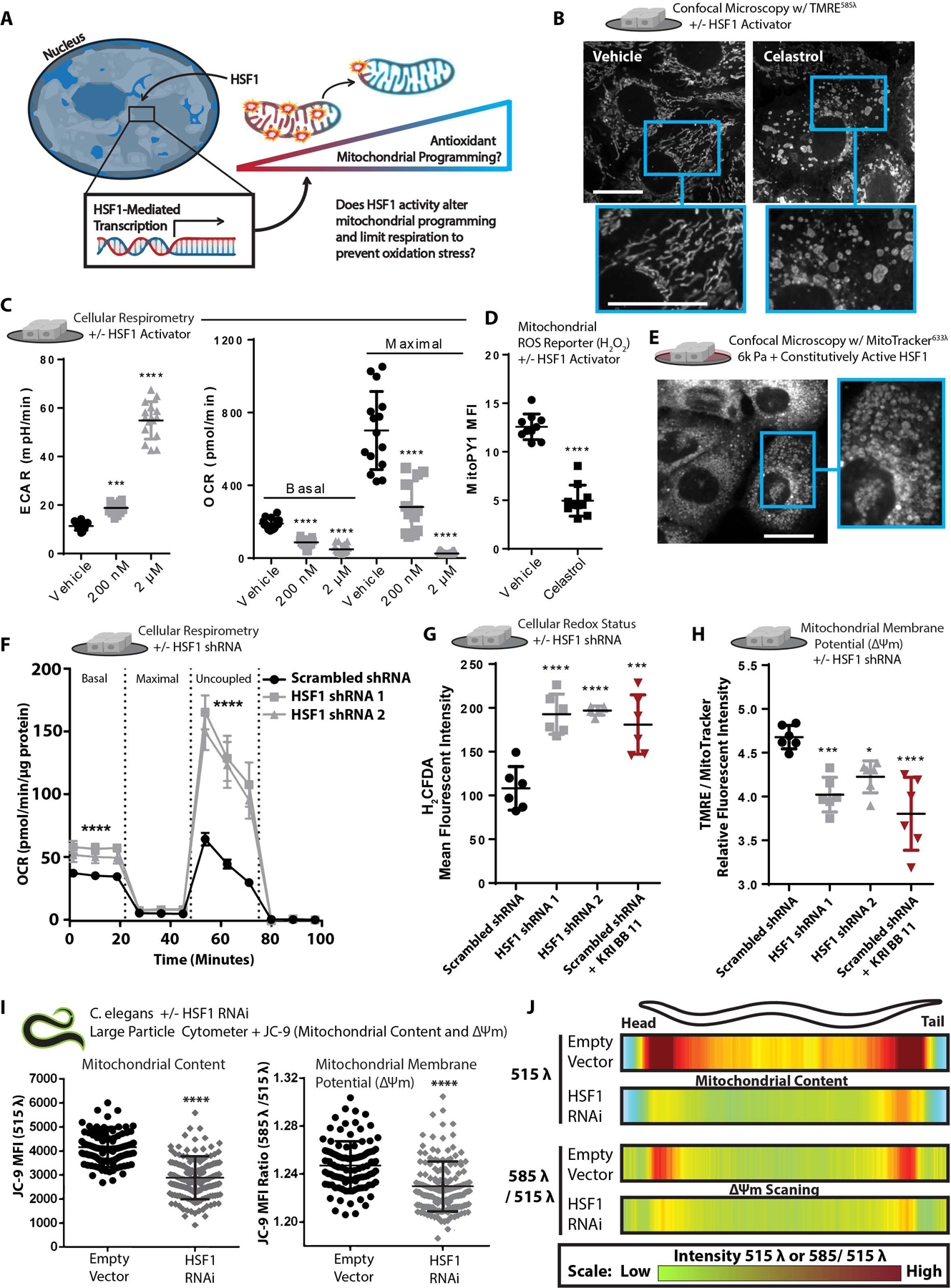
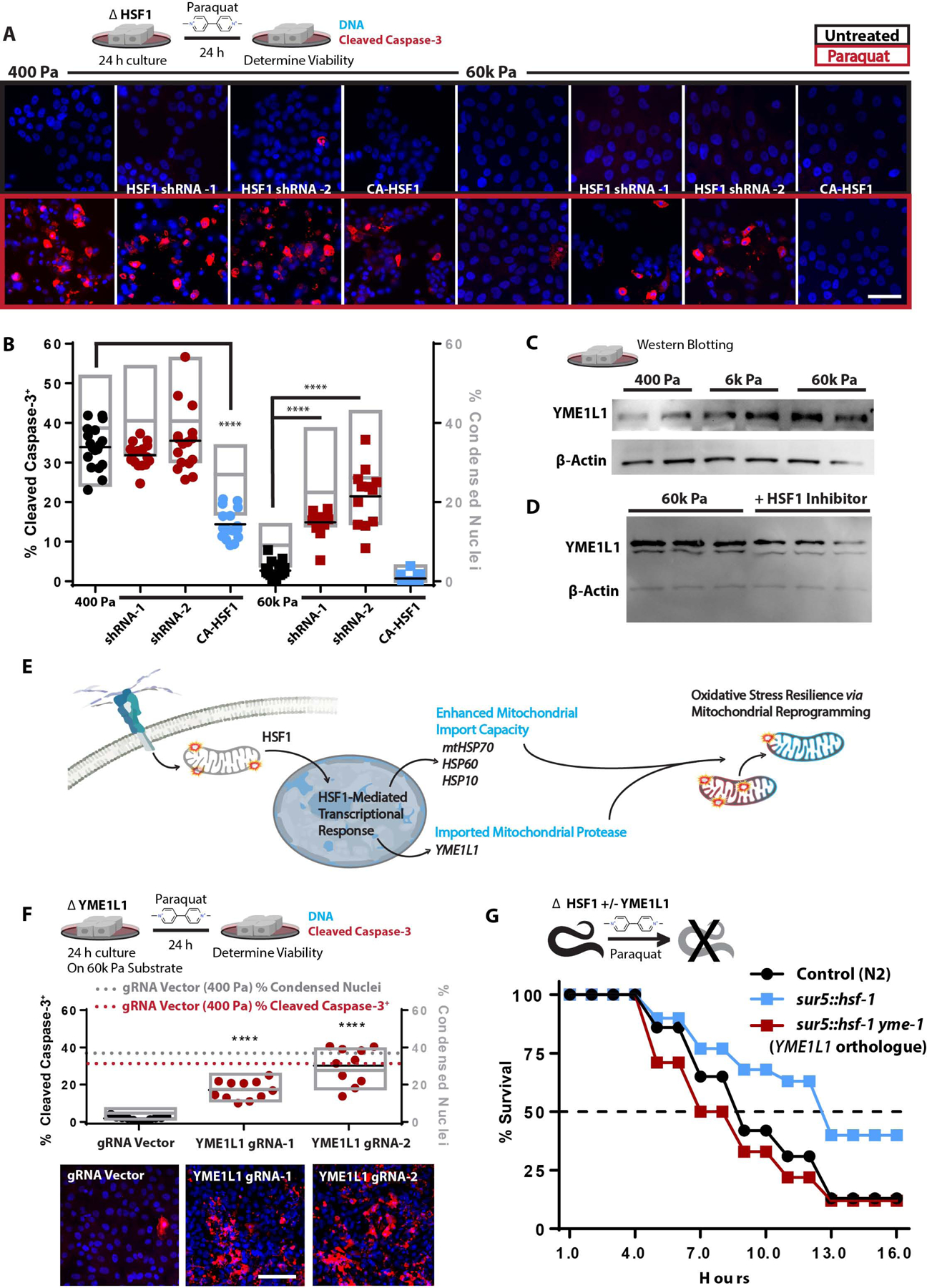
Mitochondria control eukaryotic cell fate by producing the energy needed to support life and the signals required to execute programed cell death. The biochemical milieu is known to affect mitochondrial function and contribute to the dysfunctional mitochondrial phenotypes implicated in cancer and the morbidities of aging. However, the physical characteristics of the extracellular matrix are also altered in cancerous and aging tissues. Here, we demonstrate that cells sense the physical properties of the extracellular matrix and activate a mitochondrial stress response that adaptively tunes mitochondrial function via solute carrier family 9 member A1-dependent ion exchange and heat shock factor 1-dependent transcription. Overall, our data indicate that adhesion-mediated mechanosignaling may play an unappreciated role in the altered mitochondrial functions observed in aging and cancer.
Keywords: UPRmt; adhesion; aging; cancer; extracellular matrix; mechanical stress; mechanotabolism; metabolism; oxidative stress; tension.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Mechanometabolism: Mitochondria promote resilience under pressure.Curr Biol. 2021 Jul 12;31(13):R859-R861. doi: 10.1016/j.cub.2021.05.065. Curr Biol. 2021. PMID: 34256921
-
Matrix stiffness primes cells for future oxidative stress.Trends Cancer. 2021 Oct;7(10):883-885. doi: 10.1016/j.trecan.2021.08.003. Epub 2021 Aug 18. Trends Cancer. 2021. PMID: 34417163
References
-
- Anesti V, and Scorrano L (2006). The relationship between mitochondrial shape and function and the cytoskeleton. Biochim. Biophys. Acta 1757, 692–699. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- S10 OD016387/OD/NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- R01 CA222508/CA/NCI NIH HHS/United States
- U54 CA210184/CA/NCI NIH HHS/United States
- RF1 AG059751/AG/NIA NIH HHS/United States
- R35 CA242447/CA/NCI NIH HHS/United States
- T32 CA108462/CA/NCI NIH HHS/United States
- F32 CA236156/CA/NCI NIH HHS/United States
- R35 GM130332/GM/NIGMS NIH HHS/United States
- R01 CA192914/CA/NCI NIH HHS/United States
- K99 AG065200/AG/NIA NIH HHS/United States
- R01 NS059690/NS/NINDS NIH HHS/United States
- R01 AG055891/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
