

Extracellular phosphate enhances the function of F508del-CFTR rescued by CFTR correctors
- PMID: 34020896
- PMCID: PMC8503924
- DOI: 10.1016/j.jcf.2021.04.013
Extracellular phosphate enhances the function of F508del-CFTR rescued by CFTR correctors
Abstract
Background: The clinical response to cystic fibrosis transmembrane conductance regulator (CFTR) modulators varies between people with cystic fibrosis (CF) of the same genotype, in part through the action of solute carriers encoded by modifier genes. Here, we investigate whether phosphate transport by SLC34A2 modulates the function of F508del-CFTR after its rescue by CFTR correctors.
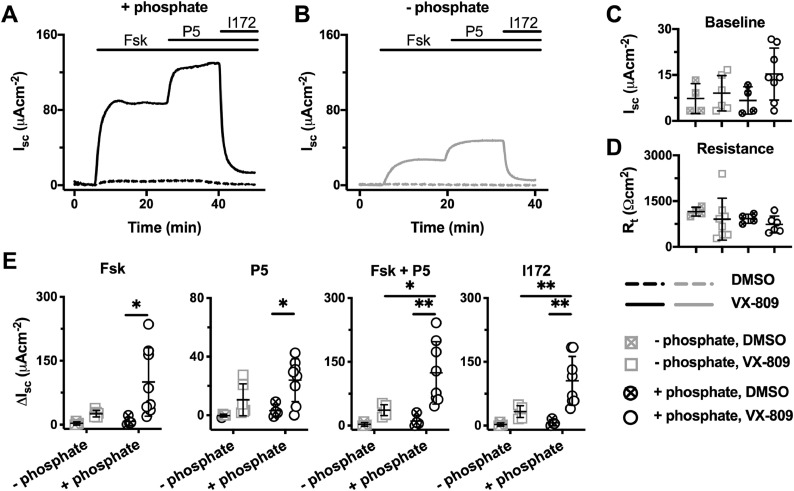
Methods: With Fischer rat thyroid (FRT) cells heterologously expressing wild-type and F508del-CFTR and fully-differentiated CF and non-CF human airway epithelial cells, we studied SLC34A2 expression and the effects of phosphate on CFTR-mediated transepithelial ion transport. F508del-CFTR was trafficked to the plasma membrane by incubation with different CFTR correctors (alone or in combination) or by low temperature.
Results: Quantitative RT-PCR demonstrated that both FRT and primary airway epithelial cells express SLC34A2 mRNA and no differences were found between cells expressing wild-type and F508del-CFTR. For both heterologously expressed and native F508del-CFTR rescued by either VX-809 or C18, the magnitude of CFTR-mediated Cl- currents was dependent on the presence of extracellular phosphate. However, this effect of phosphate was not detected with wild-type and low temperature-rescued F508del-CFTR Cl- currents. Importantly, the modulatory effect of phosphate was observed in native CF airway cells exposed to VX-445, VX-661 and VX-770 (Trikafta) and was dependent on the presence of both sodium and phosphate.
Conclusions: Extracellular phosphate modulates the magnitude of CFTR-mediated Cl- currents after F508del-CFTR rescue by clinically-approved CFTR correctors. This effect likely involves electrogenic phosphate transport by SLC34A2. It might contribute to inter-individual variability in the clinical response to CFTR correctors.
Keywords: Airway epithelia; CFTR correctors; Elexacaftor-tezacaftor-ivacaftor (Trikafta); F508del-CFTR; Phosphate; SLC34A2.
Copyright © 2021 The Author(s). Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest The authors declare that they have no conflicts of interest with the contents of this manuscript.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
