

Cutaneous innate immune tolerance is mediated by epigenetic control of MAP2K3 by HDAC8/9
- PMID: 34021025
- PMCID: PMC8363943
- DOI: 10.1126/sciimmunol.abe1935
Cutaneous innate immune tolerance is mediated by epigenetic control of MAP2K3 by HDAC8/9
Abstract
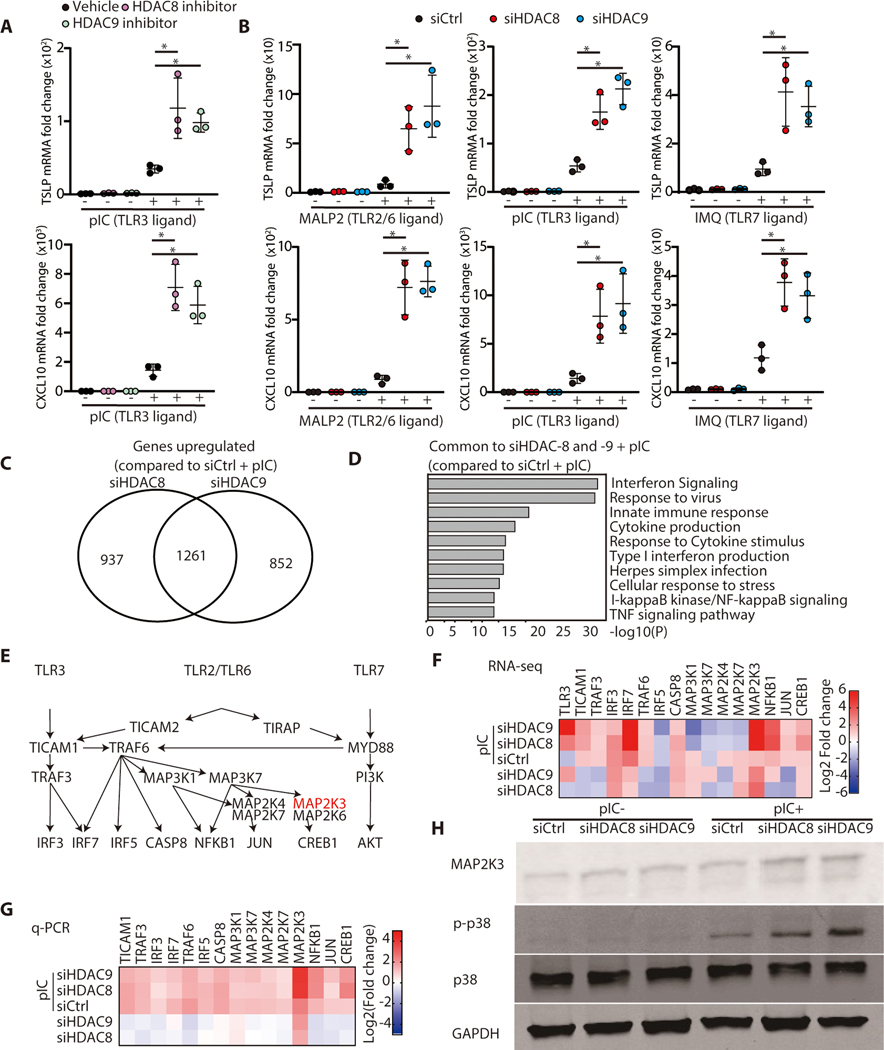
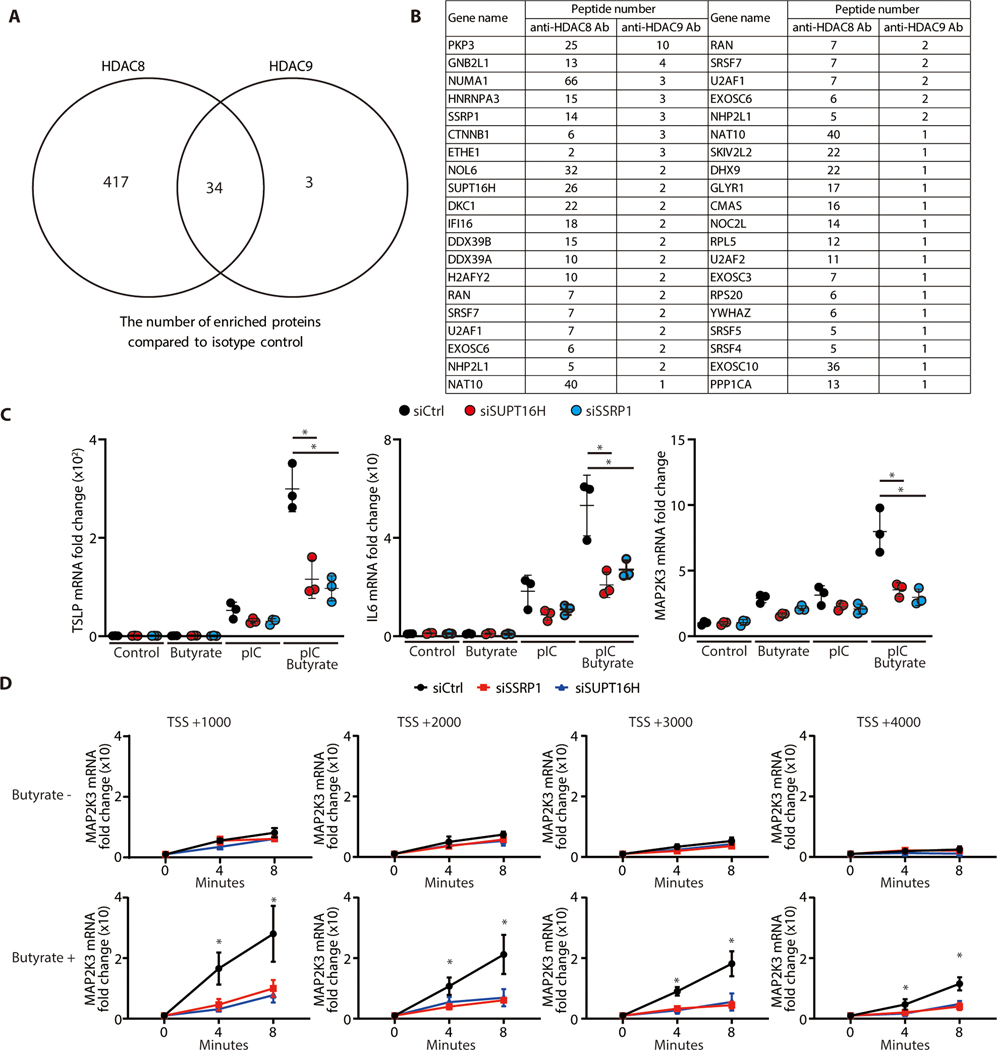
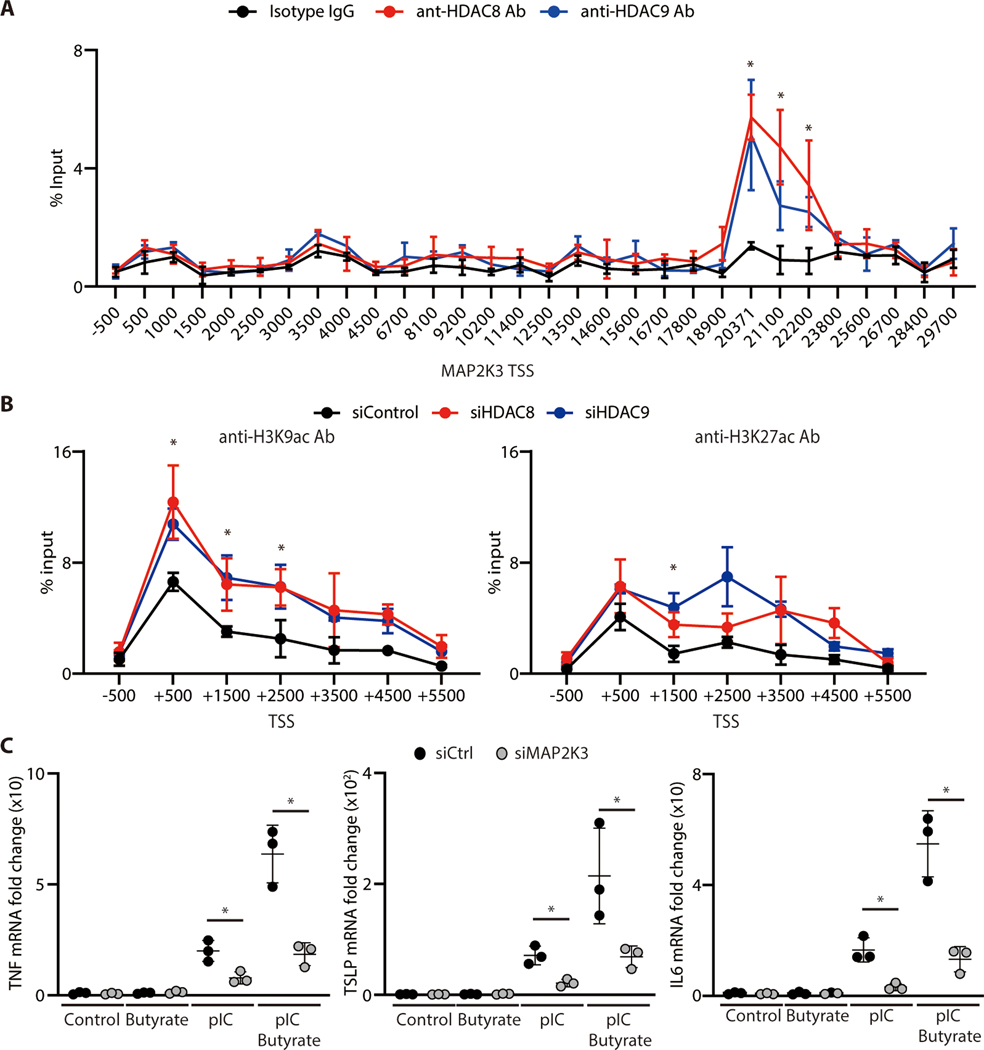
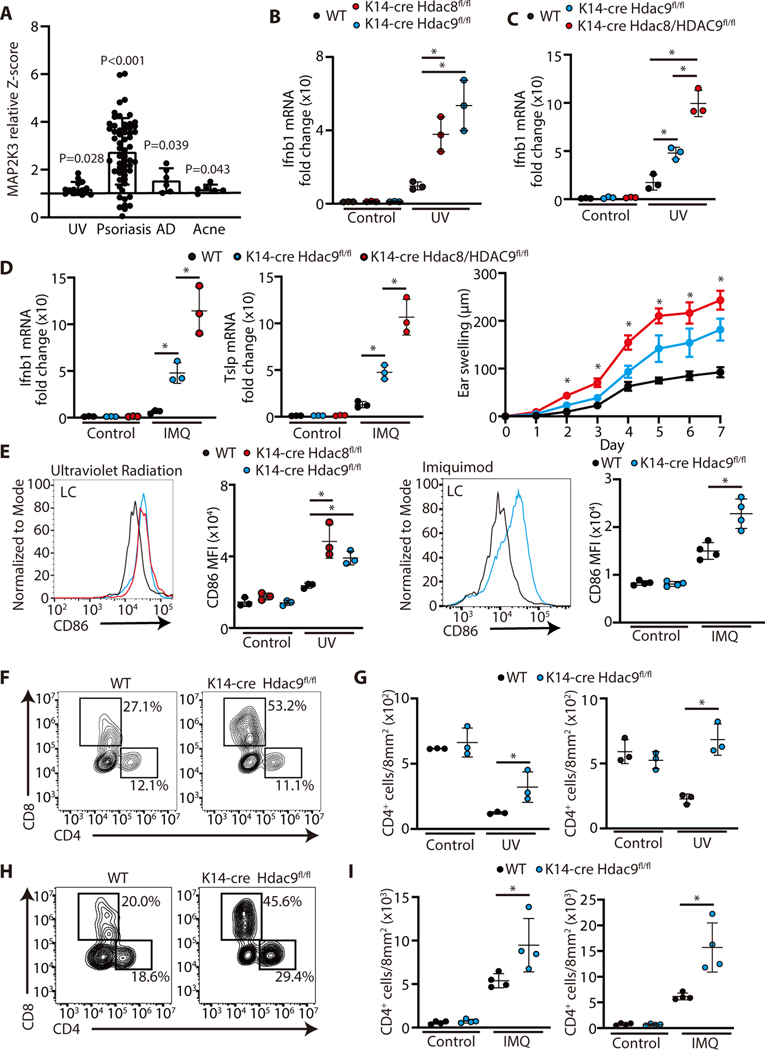
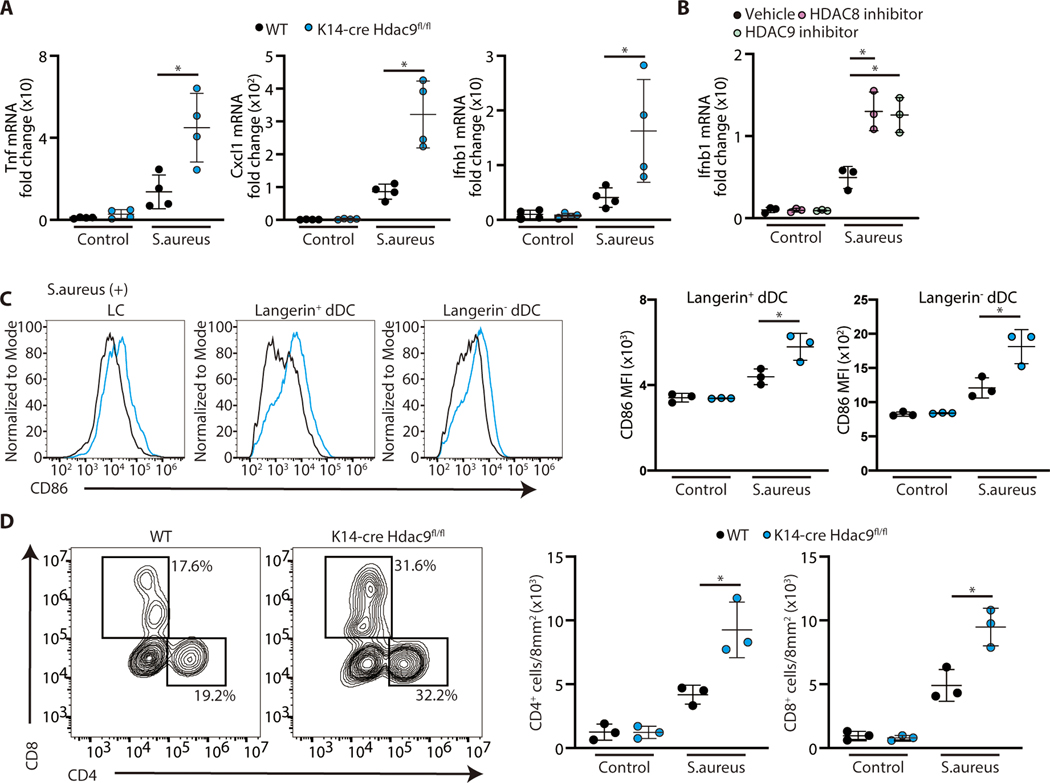
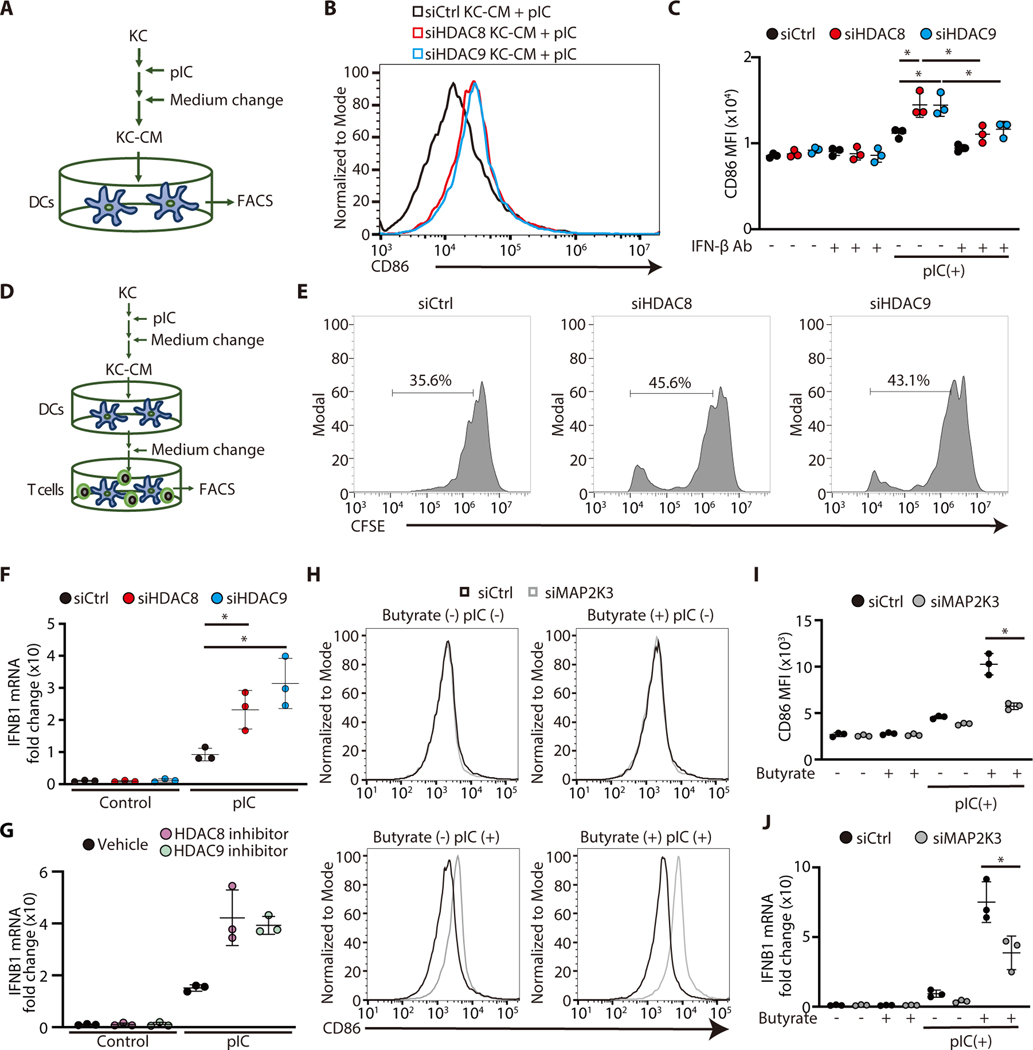
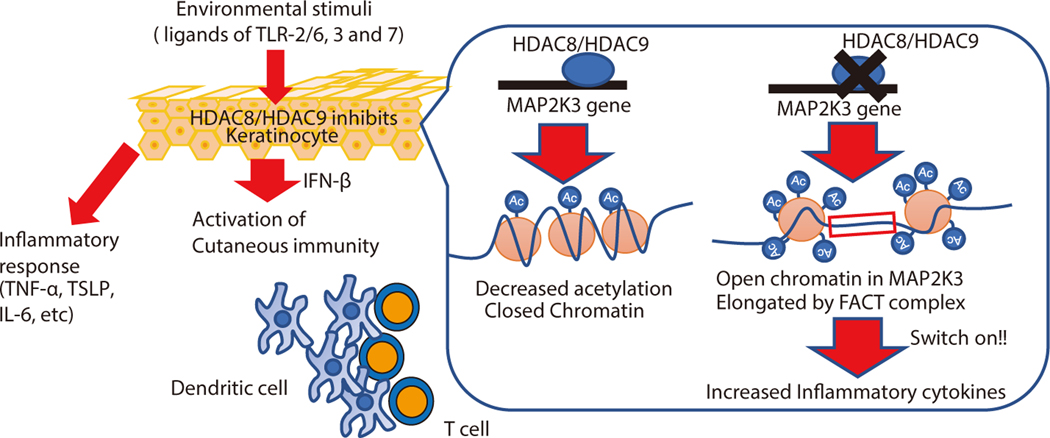
The skin typically tolerates exposure to various microbes and chemicals in the environment. Here, we investigated how the epidermis maintains this innate immune tolerance to stimuli that are recognized by Toll-like receptors (TLRs). Loss of tolerance to TLR ligands occurred after silencing of the histone deacetylases (HDACs) HDAC8 and HDAC9 in keratinocytes. Transcriptional analysis identified MAP2K3 as suppressed by HDAC8/9 activity and a potential key intermediary for establishing this tolerance. HDAC8/9 influenced acetylation at H3K9 and H3K27 marks in the MAP2K3 promoter. Proteomic analysis further identified SSRP1 and SUPT16H as associated with HDAC8/9 and responsible for transcriptional elongation of MAP2K3. Silencing of MAP2K3 blocked the capacity of HDAC8/9 to influence cytokine responses. Relevance in vivo was supported by observations of increased MAP2K3 in human inflammatory skin conditions and the capacity of keratinocyte HDAC8/9 to influence dendritic cell maturation and T cell proliferation. Keratinocyte-specific deletion of HDAC8/9 also increased inflammation in mice after exposure to ultraviolet radiation, imiquimod, or Staphylococcus aureus These findings define a mechanism for the epidermis to regulate inflammation in the presence of ubiquitous TLR ligands.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
References
-
- Dainichi T. et al., The epithelial immune microenvironment (EIME) in atopic dermatitis and psoriasis. Nat Immunol 19, 1286–1298 (2018). - PubMed
-
- Genuis SJ, Sensitivity-related illness: the escalating pandemic of allergy, food intolerance and chemical sensitivity. Sci Total Environ 408, 6047–6061 (2010). - PubMed
-
- Alferink J. et al., Control of neonatal tolerance to tissue antigens by peripheral T cell trafficking. Science 282, 1338–1341 (1998). - PubMed
-
- Zhang X, Zhivaki D, Lo-Man R, Unique aspects of the perinatal immune system. Nat Rev Immunol 17, 495–507 (2017). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
