

Defining the Energetic Basis for a Conformational Switch Mediating Ligand-Independent Activation of Mutant Estrogen Receptors in Breast Cancer
- PMID: 34021071
- PMCID: PMC8419021
- DOI: 10.1158/1541-7786.MCR-20-1017
Defining the Energetic Basis for a Conformational Switch Mediating Ligand-Independent Activation of Mutant Estrogen Receptors in Breast Cancer
Abstract
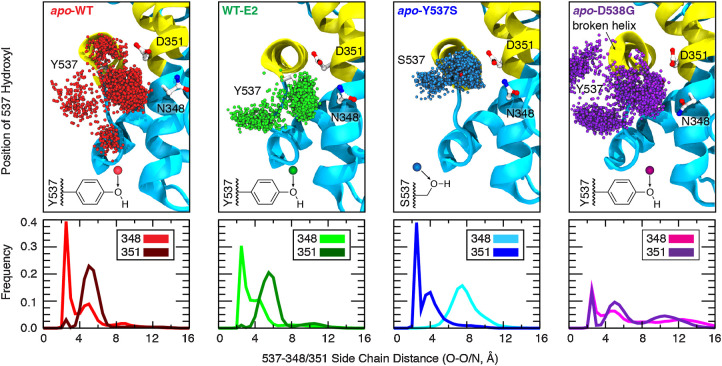
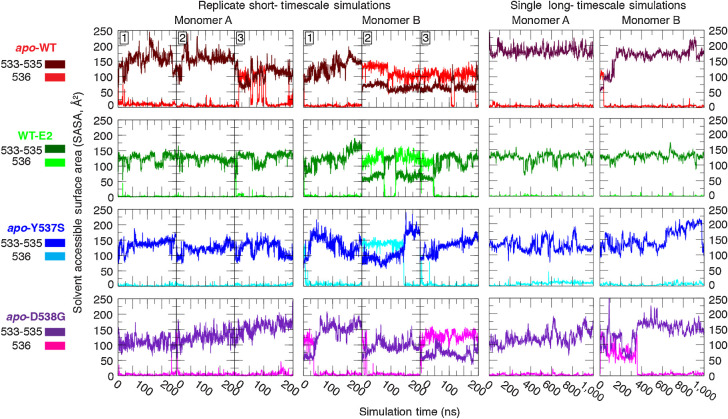
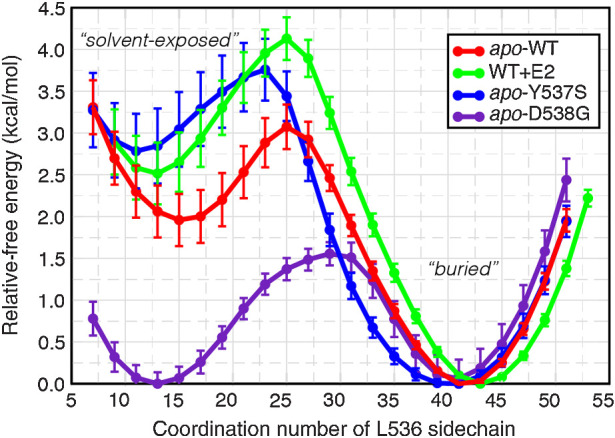
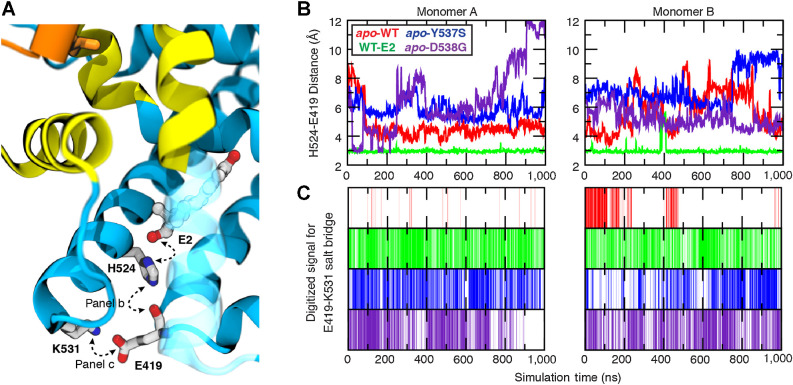
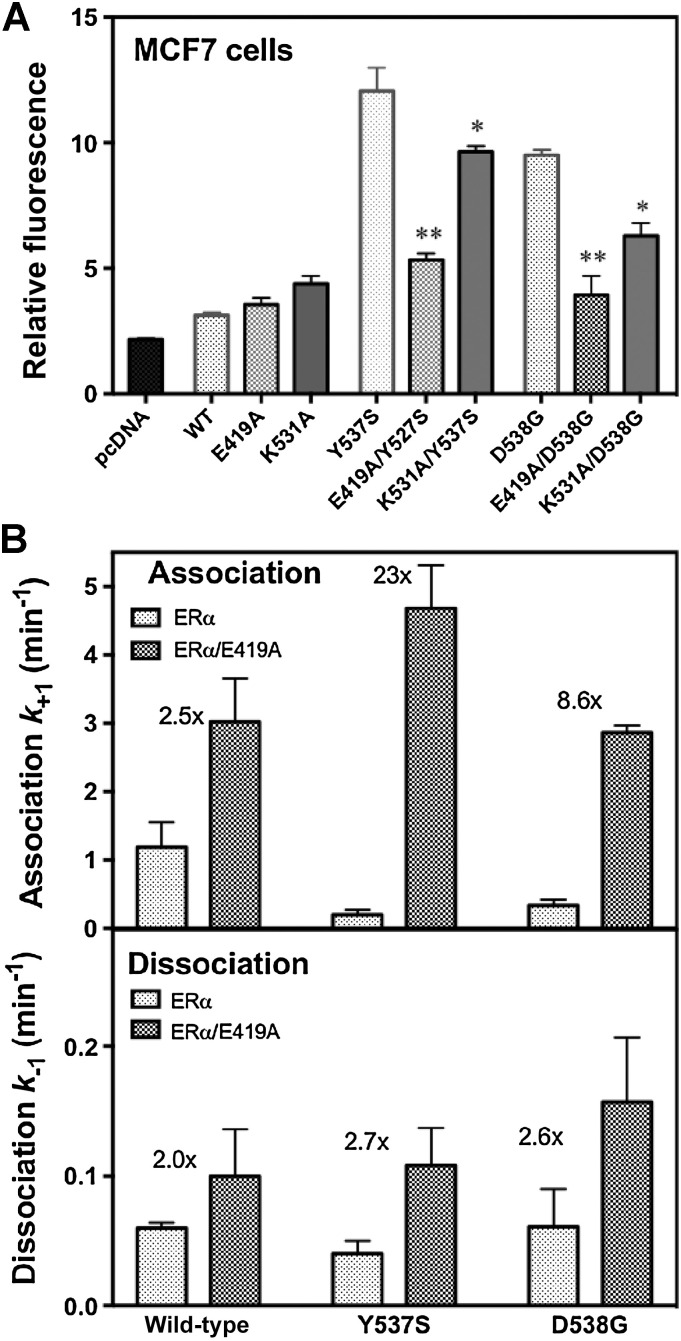
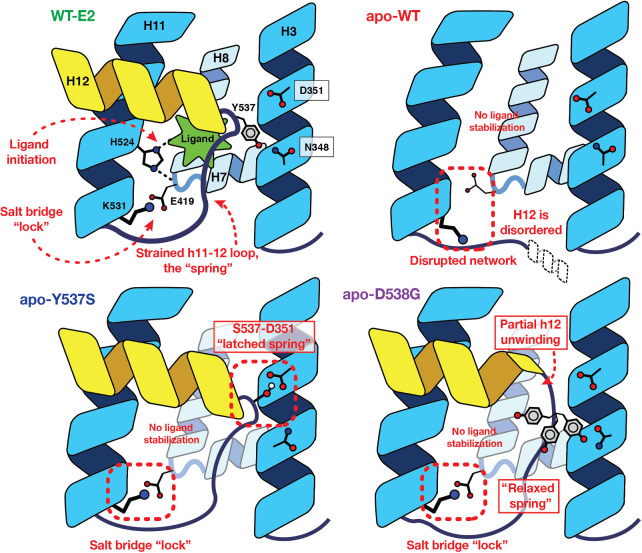
Although most primary estrogen receptor (ER)-positive breast cancers respond well to endocrine therapies, many relapse later as metastatic disease due to endocrine therapy resistance. Over one third of these are associated with mutations in the ligand-binding domain (LBD) that activate the receptor independent of ligand. We have used an array of advanced computational techniques rooted in molecular dynamics simulations, in concert with and validated by experiments, to characterize the molecular mechanisms by which specific acquired somatic point mutations give rise to ER constitutive activation. By comparing structural and energetic features of constitutively active mutants and ligand-bound forms of ER-LBD with unliganded wild-type (WT) ER, we characterize a spring force originating from strain in the Helix 11-12 loop of WT-ER, opposing folding of Helix 12 into the active conformation and keeping WT-ER off and disordered, with the ligand-binding pocket open for rapid ligand binding. We quantify ways in which this spring force is abrogated by activating mutations that latch (Y537S) or relax (D538G) the folded form of the loop, enabling formation of the active conformation without ligand binding. We also identify a new ligand-mediated hydrogen-bonding network that stabilizes the active, ligand-bound conformation of WT-ER LBD, and similarly stabilizes the active conformation of the ER mutants in the hormone-free state. IMPLICATIONS: Our investigations provide deep insight into the energetic basis for the structural mechanisms of receptor activation through mutation, exemplified here with ER in endocrine-resistant metastatic breast cancers, with potential application to other dysregulated receptor signaling due to driver mutations.
©2021 The Authors; Published by the American Association for Cancer Research.
Figures
Similar articles
-
Lasofoxifene as a potential treatment for therapy-resistant ER-positive metastatic breast cancer.Breast Cancer Res. 2021 May 12;23(1):54. doi: 10.1186/s13058-021-01431-w. Breast Cancer Res. 2021. PMID: 33980285 Free PMC article.
-
Altered ligand binding properties and enhanced stability of a constitutively active estrogen receptor: evidence that an open pocket conformation is required for ligand interaction.Biochemistry. 1997 Dec 2;36(48):14897-905. doi: 10.1021/bi971746l. Biochemistry. 1997. PMID: 9398213
-
Constitutively active human estrogen receptors containing amino acid substitutions for tyrosine 537 in the receptor protein.Mol Endocrinol. 1996 Nov;10(11):1388-98. doi: 10.1210/mend.10.11.8923465. Mol Endocrinol. 1996. PMID: 8923465
-
A Closer Look at Estrogen Receptor Mutations in Breast Cancer and Their Implications for Estrogen and Antiestrogen Responses.Int J Mol Sci. 2021 Jan 13;22(2):756. doi: 10.3390/ijms22020756. Int J Mol Sci. 2021. PMID: 33451133 Free PMC article. Review.
-
Structural and functional characterization of aromatase, estrogen receptor, and their genes in endocrine-responsive and -resistant breast cancer cells.J Steroid Biochem Mol Biol. 2016 Jul;161:73-83. doi: 10.1016/j.jsbmb.2015.07.018. Epub 2015 Aug 13. J Steroid Biochem Mol Biol. 2016. PMID: 26277097 Free PMC article. Review.
Cited by
-
Recent Applications of In Silico Approaches for Studying Receptor Mutations Associated with Human Pathologies.Molecules. 2024 Nov 13;29(22):5349. doi: 10.3390/molecules29225349. Molecules. 2024. PMID: 39598735 Free PMC article. Review.
-
Membrane-bound model of the ternary complex between factor VIIa/tissue factor and factor X.Blood Adv. 2025 Feb 25;9(4):729-740. doi: 10.1182/bloodadvances.2024014845. Blood Adv. 2025. PMID: 39671302 Free PMC article.
-
Evolutionary Overview and Future Perspectives: ESR1 Mutations, Liquid Biopsy, and Artificial Intelligence for a New Era of Personalized Medicine in ER+ Breast Cancer.Mol Diagn Ther. 2025 Aug 30. doi: 10.1007/s40291-025-00811-8. Online ahead of print. Mol Diagn Ther. 2025. PMID: 40884685 Review.
-
Somatic estrogen receptor α mutations that induce dimerization promote receptor activity and breast cancer proliferation.J Clin Invest. 2024 Jan 2;134(1):e163242. doi: 10.1172/JCI163242. J Clin Invest. 2024. PMID: 37883178 Free PMC article.
-
Estrogen Receptor Alpha Mutations, Truncations, Heterodimers, and Therapies.Endocrinology. 2024 Apr 29;165(6):bqae051. doi: 10.1210/endocr/bqae051. Endocrinology. 2024. PMID: 38643482 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
