

In vivo tissue-specific chromatin profiling in Drosophila melanogaster using GFP-tagged nuclei
- PMID: 34022041
- PMCID: PMC8864739
- DOI: 10.1093/genetics/iyab079
In vivo tissue-specific chromatin profiling in Drosophila melanogaster using GFP-tagged nuclei
Abstract
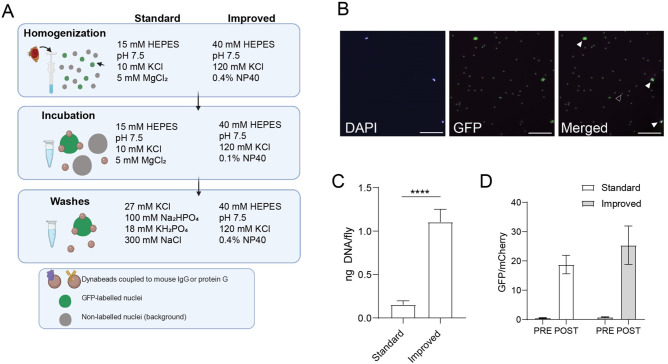
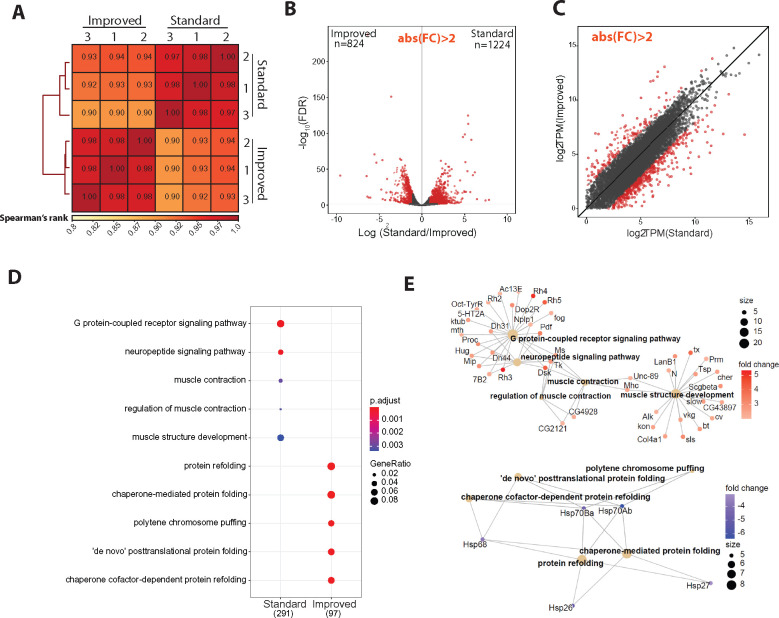
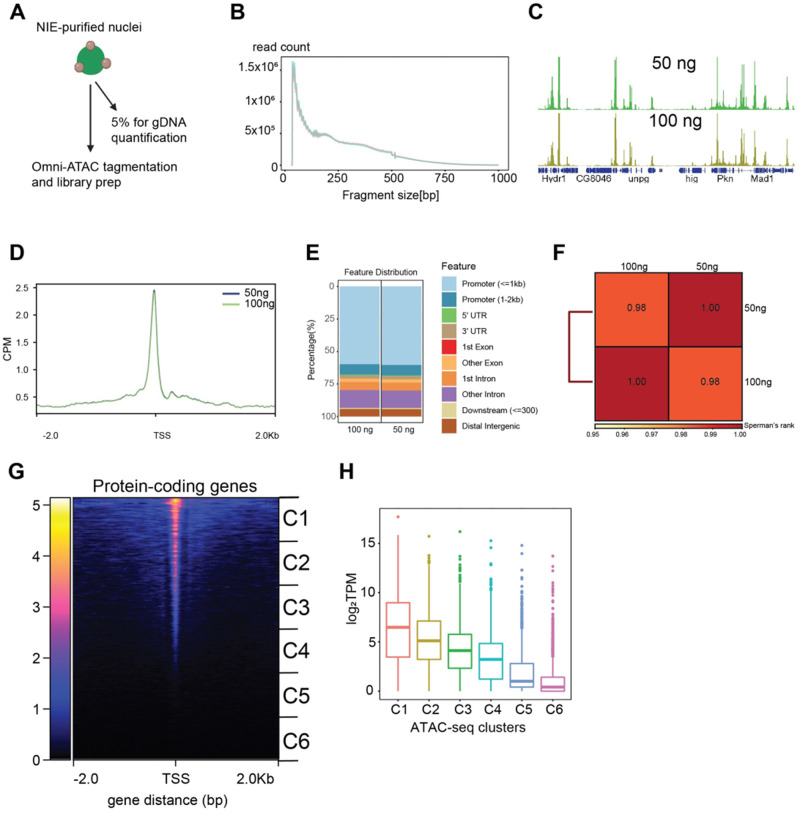
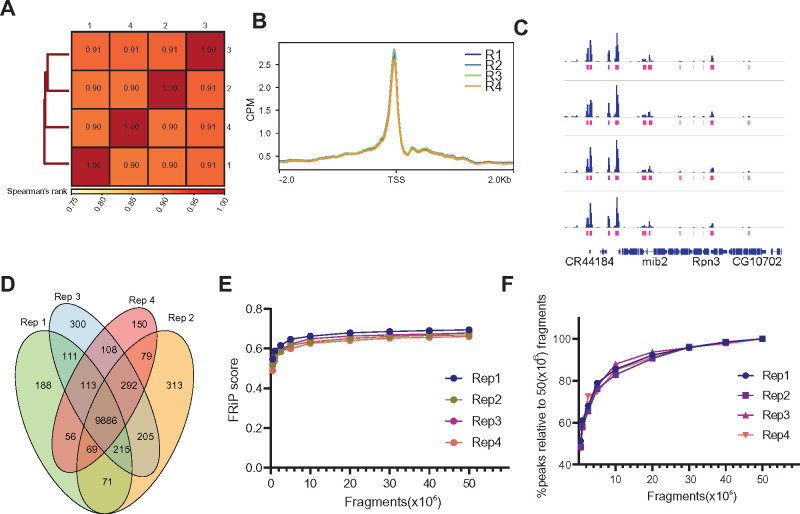
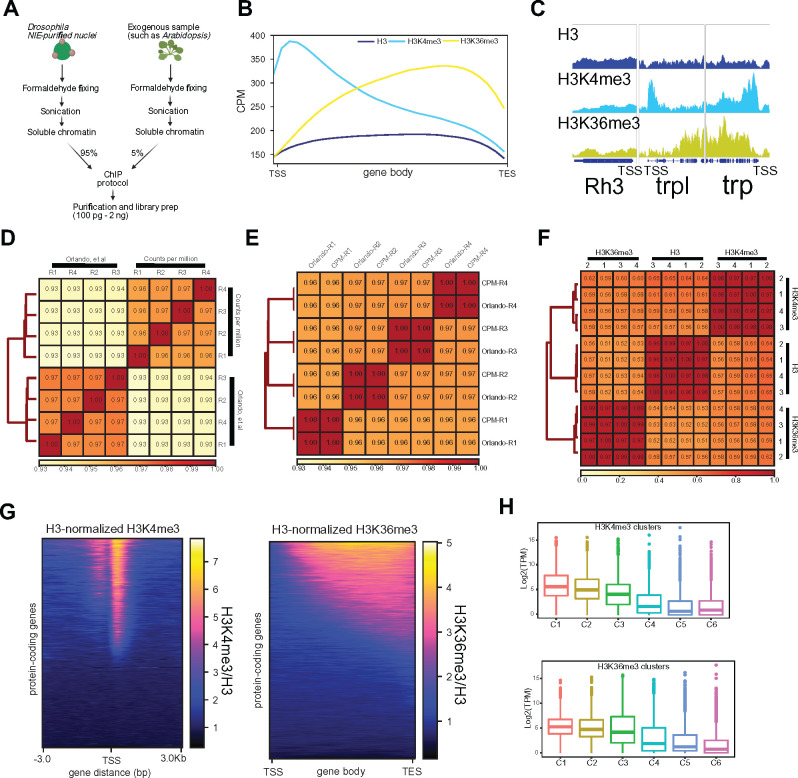
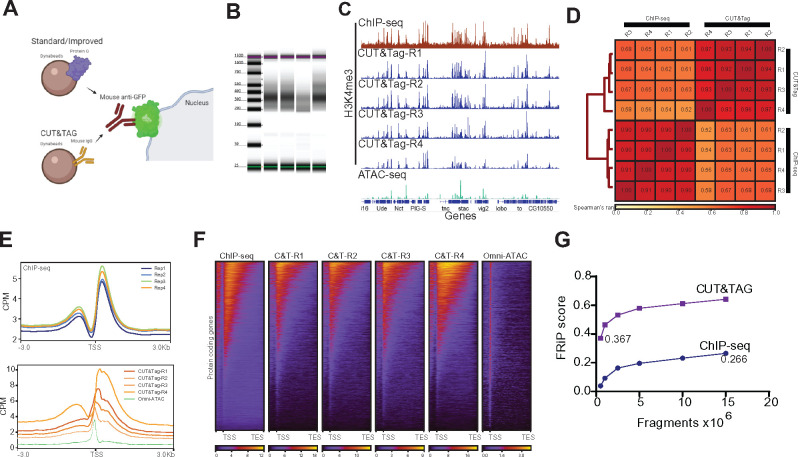
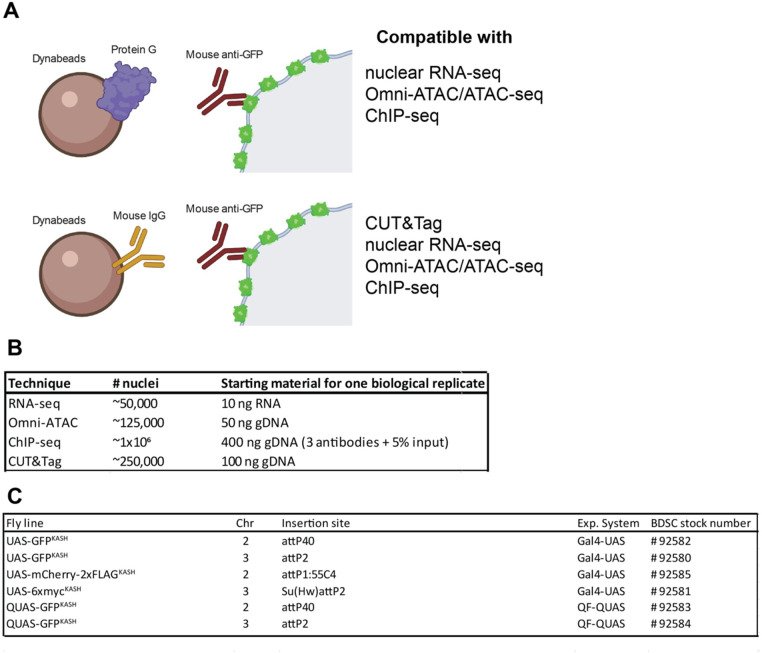
The chromatin landscape defines cellular identity in multicellular organisms with unique patterns of DNA accessibility and histone marks decorating the genome of each cell type. Thus, profiling the chromatin state of different cell types in an intact organism under disease or physiological conditions can provide insight into how chromatin regulates cell homeostasis in vivo. To overcome the many challenges associated with characterizing chromatin state in specific cell types, we developed an improved approach to isolate Drosophila melanogaster nuclei tagged with a GFPKASH protein. The perinuclear space-localized KASH domain anchors GFP to the outer nuclear membrane, and expression of UAS-GFPKASH can be controlled by tissue-specific Gal4 drivers. Using this protocol, we profiled chromatin accessibility using an improved version of Assay for Transposable Accessible Chromatin followed by sequencing (ATAC-seq), called Omni-ATAC. In addition, we examined the distribution of histone marks using Chromatin immunoprecipitation followed by sequencing (ChIP-seq) and Cleavage Under Targets and Tagmentation (CUT&Tag) in adult photoreceptor neurons. We show that the chromatin landscape of photoreceptors reflects the transcriptional state of these cells, demonstrating the quality and reproducibility of our approach for profiling the transcriptome and epigenome of specific cell types in Drosophila.
Keywords: Drosophila; ATAC-seq; cell type; chromatin; photoreceptor; transcription.
© The Author(s) 2021. Published by Oxford University Press on behalf of Genetics Society of America. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures
Similar articles
-
Systematic alteration of ATAC-seq for profiling open chromatin in cryopreserved nuclei preparations from livestock tissues.Sci Rep. 2020 Mar 23;10(1):5230. doi: 10.1038/s41598-020-61678-9. Sci Rep. 2020. PMID: 32251359 Free PMC article.
-
Review and Evaluate the Bioinformatics Analysis Strategies of ATAC-seq and CUT&Tag Data.Genomics Proteomics Bioinformatics. 2024 Sep 13;22(3):qzae054. doi: 10.1093/gpbjnl/qzae054. Genomics Proteomics Bioinformatics. 2024. PMID: 39255248 Free PMC article. Review.
-
Integrative Analysis of CUT&Tag and RNA-Seq Data Through Bioinformatics: A Unified Workflow for Enhanced Insights.Methods Mol Biol. 2024;2846:191-213. doi: 10.1007/978-1-0716-4071-5_13. Methods Mol Biol. 2024. PMID: 39141238
-
Optimized assay for transposase-accessible chromatin by sequencing (ATAC-seq) library preparation from adult Drosophila melanogaster neurons.Sci Rep. 2022 Apr 11;12(1):6043. doi: 10.1038/s41598-022-09869-4. Sci Rep. 2022. PMID: 35411004 Free PMC article.
-
Methods for ChIP-seq analysis: A practical workflow and advanced applications.Methods. 2021 Mar;187:44-53. doi: 10.1016/j.ymeth.2020.03.005. Epub 2020 Mar 30. Methods. 2021. PMID: 32240773 Review.
Cited by
-
The Clock:Cycle complex is a major transcriptional regulator of Drosophila photoreceptors that protects the eye from retinal degeneration and oxidative stress.PLoS Genet. 2022 Jan 31;18(1):e1010021. doi: 10.1371/journal.pgen.1010021. eCollection 2022 Jan. PLoS Genet. 2022. PMID: 35100266 Free PMC article.
-
Chiffon triggers global histone H3 acetylation and expression of developmental genes in Drosophila embryos.J Cell Sci. 2022 Jan 15;135(2):jcs259132. doi: 10.1242/jcs.259132. Epub 2022 Jan 24. J Cell Sci. 2022. PMID: 34908116 Free PMC article.
-
Proper control of R-loop homeostasis is required for maintenance of gene expression and neuronal function during aging.Aging Cell. 2022 Feb;21(2):e13554. doi: 10.1111/acel.13554. Epub 2022 Jan 20. Aging Cell. 2022. PMID: 35048512 Free PMC article.
-
Aging and Light Stress Result in Overlapping and Unique Gene Expression Changes in Photoreceptors.Genes (Basel). 2022 Jan 29;13(2):264. doi: 10.3390/genes13020264. Genes (Basel). 2022. PMID: 35205309 Free PMC article.
-
Establishing the contribution of active histone methylation marks to the aging transcriptional landscape of Drosophila photoreceptors.Sci Rep. 2023 Mar 29;13(1):5105. doi: 10.1038/s41598-023-32273-5. Sci Rep. 2023. PMID: 36991154 Free PMC article.
References
-
- Bai L, Morozov AV.. 2010. Gene regulation by nucleosome positioning. Trends Genet. 26:476–483. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
