

Local actin dynamics couple speed and persistence in a cellular Potts model of cell migration
- PMID: 34022237
- PMCID: PMC8390880
- DOI: 10.1016/j.bpj.2021.04.036
Local actin dynamics couple speed and persistence in a cellular Potts model of cell migration
Abstract
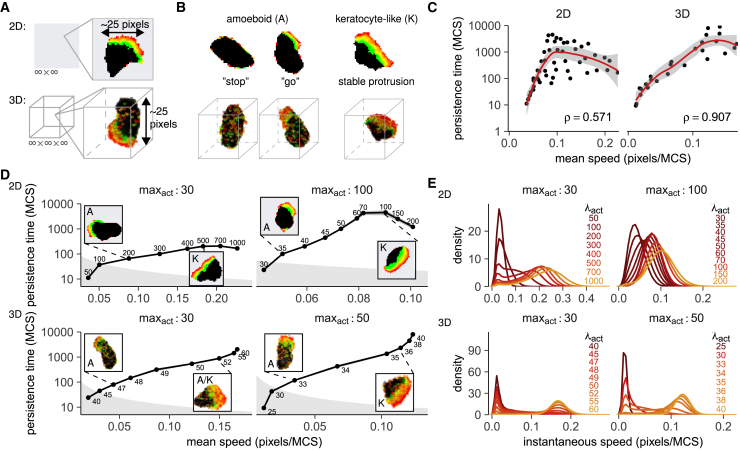
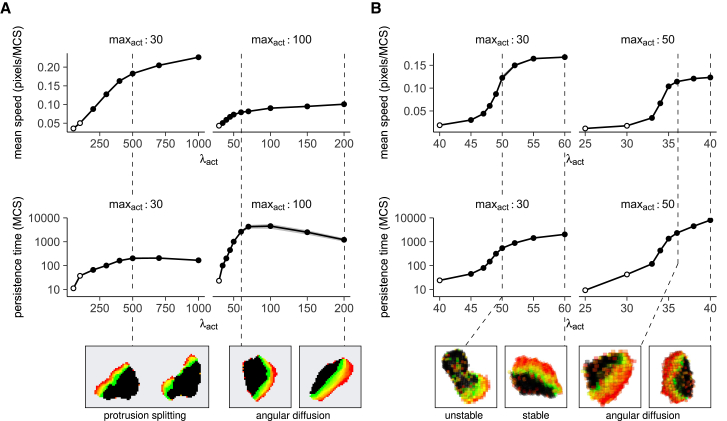
Cell migration is astoundingly diverse. Molecular signatures, cell-cell interactions, and environmental structures each play their part in shaping cell motion, yielding numerous morphologies and migration modes. Nevertheless, in recent years, a simple unifying law was found to describe cell migration across many different cell types and contexts: faster cells turn less frequently. This universal coupling between speed and persistence (UCSP) was explained by retrograde actin flow from front to back, but it remains unclear how this mechanism generalizes to cells with complex shapes and cells migrating in structured environments, which may not have a well-defined front-to-back orientation. Here, we present an in-depth characterization of an existing cellular Potts model, in which cells polarize dynamically from a combination of local actin dynamics (stimulating protrusions) and global membrane tension along the perimeter (inhibiting protrusions). We first show that the UCSP emerges spontaneously in this model through a cross talk of intracellular mechanisms, cell shape, and environmental constraints, resembling the dynamic nature of cell migration in vivo. Importantly, we find that local protrusion dynamics suffice to reproduce the UCSP-even in cases in which no clear global, front-to-back polarity exists. We then harness the spatial nature of the cellular Potts model to show how cell shape dynamics limit both the speed and persistence a cell can reach and how a rigid environment such as the skin can restrict cell motility even further. Our results broaden the range of potential mechanisms underlying the speed-persistence coupling that has emerged as a fundamental property of migrating cells.
Copyright © 2021 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures



Similar articles
-
Leading edge maintenance in migrating cells is an emergent property of branched actin network growth.Elife. 2022 Mar 11;11:e74389. doi: 10.7554/eLife.74389. Elife. 2022. PMID: 35275060 Free PMC article.
-
Actin disassembly clock determines shape and speed of lamellipodial fragments.Proc Natl Acad Sci U S A. 2011 Dec 20;108(51):20394-9. doi: 10.1073/pnas.1105333108. Epub 2011 Dec 9. Proc Natl Acad Sci U S A. 2011. PMID: 22159033 Free PMC article.
-
Actin flows mediate a universal coupling between cell speed and cell persistence.Cell. 2015 Apr 9;161(2):374-86. doi: 10.1016/j.cell.2015.01.056. Epub 2015 Mar 19. Cell. 2015. PMID: 25799384
-
Actin, microtubules and focal adhesion dynamics during cell migration.Int J Biochem Cell Biol. 2003 Jan;35(1):39-50. doi: 10.1016/s1357-2725(02)00071-7. Int J Biochem Cell Biol. 2003. PMID: 12467646 Review.
-
Cytoskeletal Crosstalk in Cell Migration.Trends Cell Biol. 2020 Sep;30(9):720-735. doi: 10.1016/j.tcb.2020.06.004. Epub 2020 Jul 13. Trends Cell Biol. 2020. PMID: 32674938 Review.
Cited by
-
Interpreting T-cell search "strategies" in the light of evolution under constraints.PLoS Comput Biol. 2023 Feb 27;19(2):e1010918. doi: 10.1371/journal.pcbi.1010918. eCollection 2023 Feb. PLoS Comput Biol. 2023. PMID: 36848395 Free PMC article.
-
Listeria motility increases the efficiency of epithelial invasion during intestinal infection.PLoS Pathog. 2022 Dec 30;18(12):e1011028. doi: 10.1371/journal.ppat.1011028. eCollection 2022 Dec. PLoS Pathog. 2022. PMID: 36584235 Free PMC article.
-
SEM2: Introducing mechanics in cell and tissue modeling using coarse-grained homogeneous particle dynamics.APL Bioeng. 2023 Dec 5;7(4):046118. doi: 10.1063/5.0166829. eCollection 2023 Dec. APL Bioeng. 2023. PMID: 38075209 Free PMC article.
-
Crawling, waving, inch worming, dilating, and pivoting mechanics of migrating cells: Lessons from Ken Jacobson.Biophys J. 2023 Sep 19;122(18):3551-3559. doi: 10.1016/j.bpj.2023.03.023. Epub 2023 Mar 17. Biophys J. 2023. PMID: 36934300 Free PMC article. Review.
-
Computational modelling of cell motility modes emerging from cell-matrix adhesion dynamics.PLoS Comput Biol. 2022 Feb 14;18(2):e1009156. doi: 10.1371/journal.pcbi.1009156. eCollection 2022 Feb. PLoS Comput Biol. 2022. PMID: 35157694 Free PMC article.
References
-
- Theilgaard-Mönch K., Knudsen S., Borregaard N. The transcriptional activation program of human neutrophils in skin lesions supports their important role in wound healing. J. Immunol. 2004;172:7684–7693. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
