

Single-cell RNA sequencing of human liver reveals hepatic stellate cell heterogeneity
- PMID: 34027339
- PMCID: PMC8121977
- DOI: 10.1016/j.jhepr.2021.100278
Single-cell RNA sequencing of human liver reveals hepatic stellate cell heterogeneity
Abstract
Background & aims: The multiple vital functions of the human liver are performed by highly specialised parenchymal and non-parenchymal cells organised in complex collaborative sinusoidal units. Although crucial for homeostasis, the cellular make-up of the human liver remains to be fully elucidated. Here, single-cell RNA-sequencing was used to unravel the heterogeneity of human liver cells, in particular of hepatocytes (HEPs) and hepatic stellate cells (HSCs).
Method: The transcriptome of ~25,000 freshly isolated human liver cells was profiled using droplet-based RNA-sequencing. Recently published data sets and RNA in situ hybridisation were integrated to validate and locate newly identified cell populations.
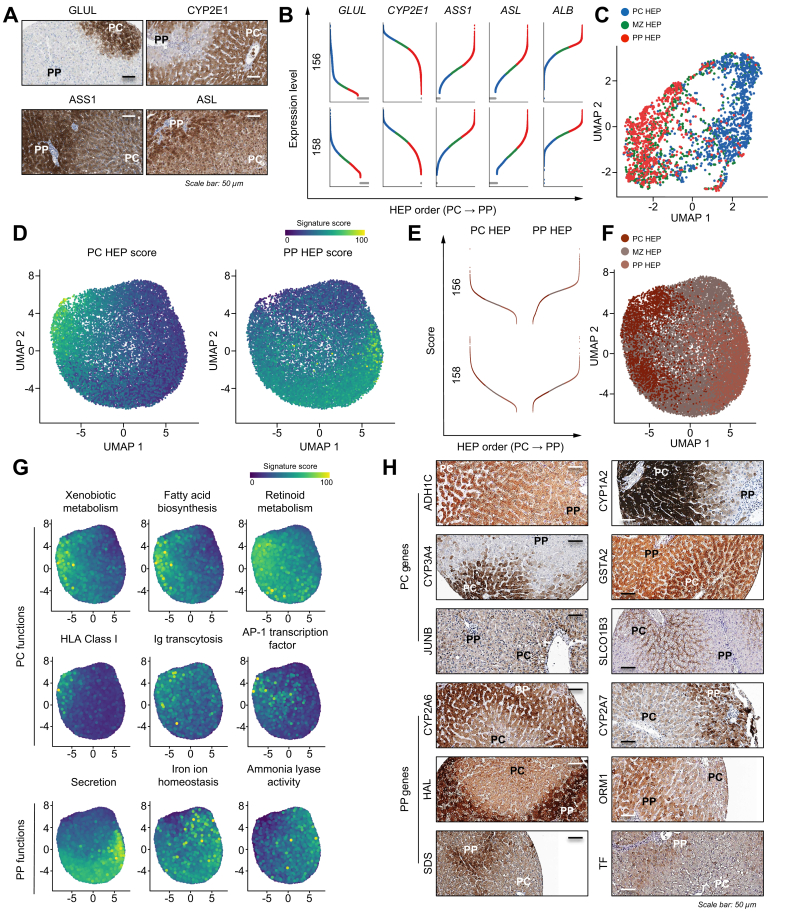
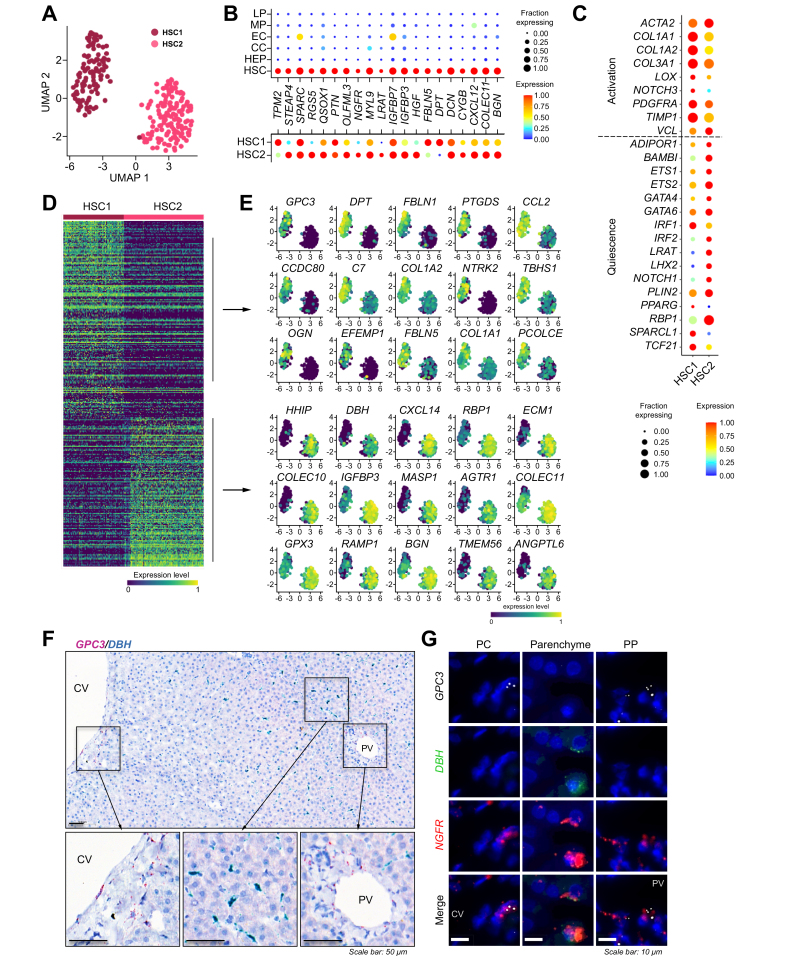
Results: In total, 22 cell populations were annotated that reflected the heterogeneity of human parenchymal and non-parenchymal liver cells. More than 20,000 HEPs were ordered along the portocentral axis to confirm known, and reveal previously undescribed, zonated liver functions. The existence of 2 subpopulations of human HSCs with unique gene expression signatures and distinct intralobular localisation was revealed (i.e. portal and central vein-concentrated GPC3 + HSCs and perisinusoidally located DBH + HSCs). In particular, these data suggest that, although both subpopulations collaborate in the production and organisation of extracellular matrix, GPC3 + HSCs specifically express genes involved in the metabolism of glycosaminoglycans, whereas DBH + HSCs display a gene signature that is reminiscent of antigen-presenting cells.
Conclusions: This study highlights metabolic zonation as a key determinant of HEP transcriptomic heterogeneity and, for the first time, outlines the existence of heterogeneous HSC subpopulations in the human liver. These findings call for further research on the functional implications of liver cell heterogeneity in health and disease.
Lay summary: This study resolves the cellular landscape of the human liver in an unbiased manner and at high resolution to provide new insights into human liver cell biology. The results highlight the physiological heterogeneity of human hepatic stellate cells.
Keywords: BSA, bovine serum albumin; CC, cholangiocyte; CV, central vein; DEG, differentially expressed gene; EC, endothelial cell; ECM, extracellular matrix; Extracellular matrix; FFPE, formaldehyde-fixed paraffin embedded; GAG, glycosaminoglycan; GEO, Gene Expression Omnibus; GO, gene ontology; HEP, hepatocyte; HLA, human leukocyte antigen; HRP, horseradish peroxidase; HSC, hepatic stellate cell; Hepatocyte; ISH, in situ hybridisation; KLR, killer lectin-like receptor; LP, lymphoid cell; Liver cell atlas; MP, macrophage; MZ, midzonal; PC, pericentral; PP, periportal; PV, portal vein; TBS, Tris buffered saline; TSA, tyramide signal amplification; UMAP, uniform manifold approximation and projection; UMI, unique molecular identifier; VIM, vimentin; Zonation; scRNA-seq, single-cell RNA-sequencing.
© 2021 Published by Elsevier B.V. on behalf of European Association for the Study of the Liver (EASL).
Conflict of interest statement
The authors declare no conflicts of interest that pertain to this work. Please refer to the accompanying ICMJE disclosure forms for further details.
Figures



References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
