

Midbrain dopaminergic inputs gate amygdala intercalated cell clusters by distinct and cooperative mechanisms in male mice
- PMID: 34028352
- PMCID: PMC8143799
- DOI: 10.7554/eLife.63708
Midbrain dopaminergic inputs gate amygdala intercalated cell clusters by distinct and cooperative mechanisms in male mice
Abstract
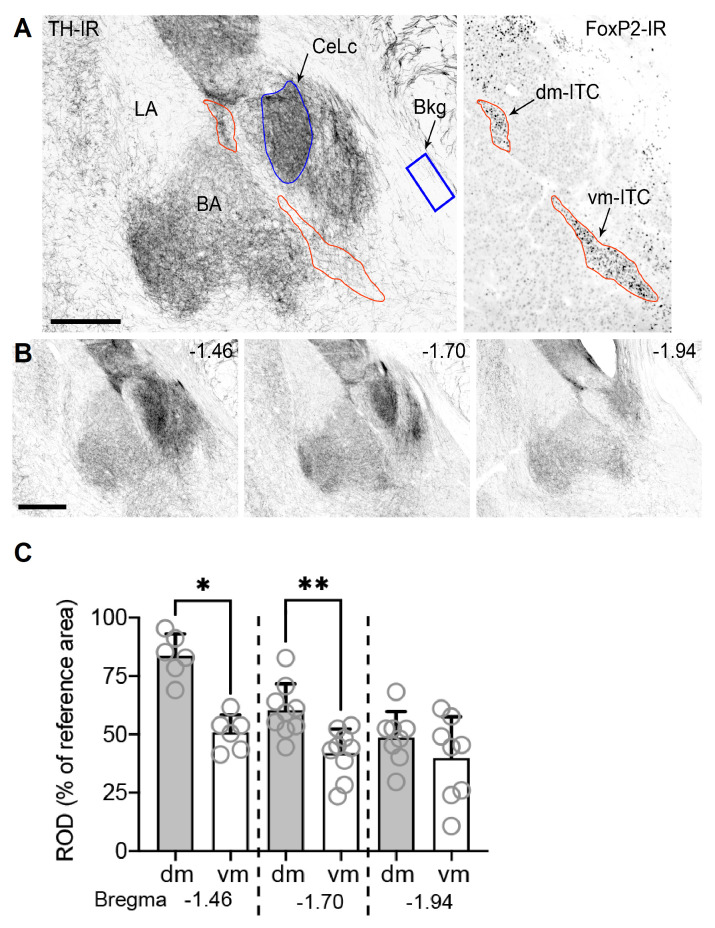
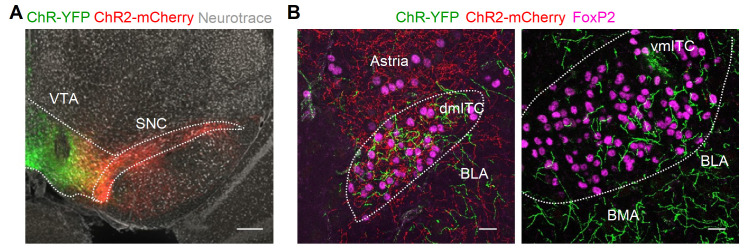
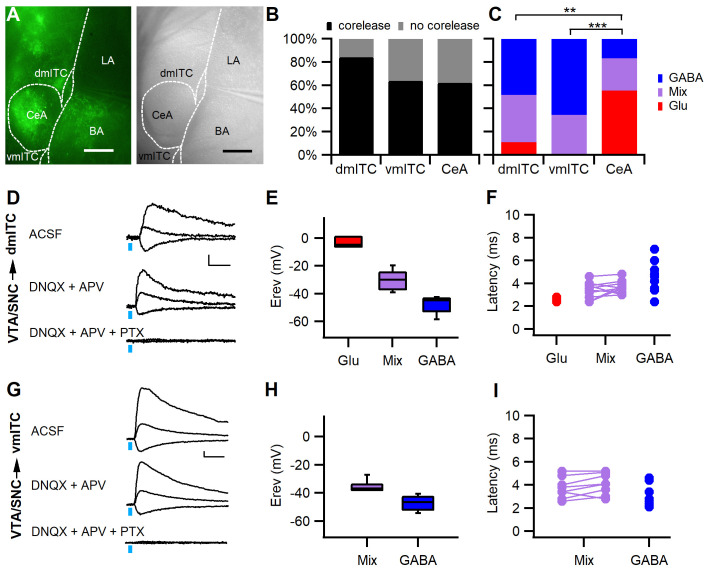
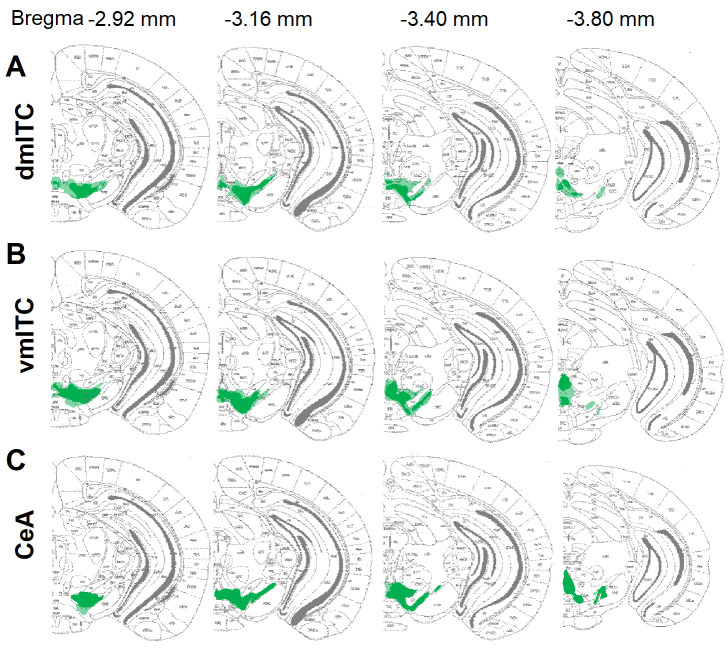
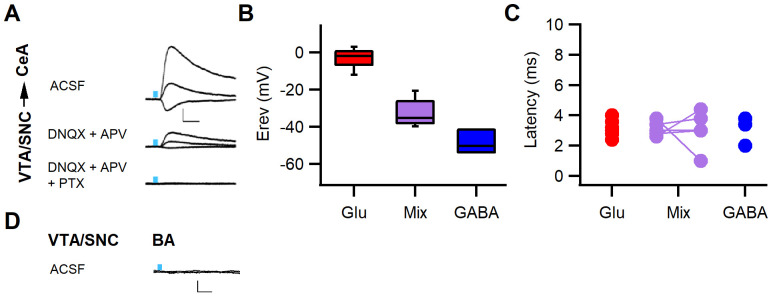
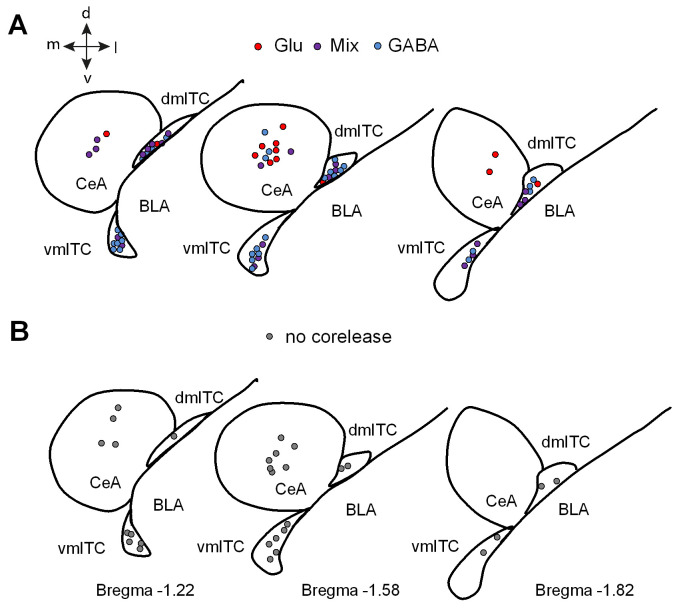
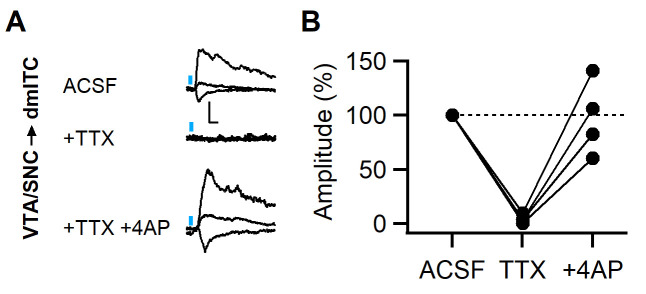
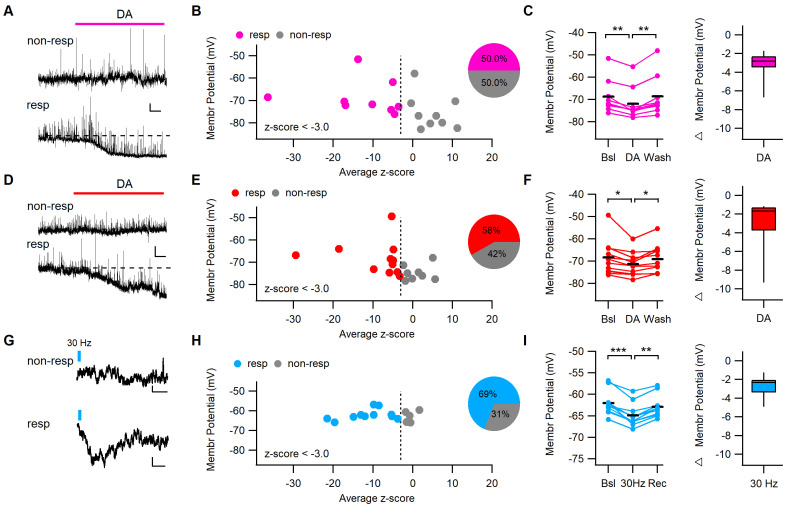
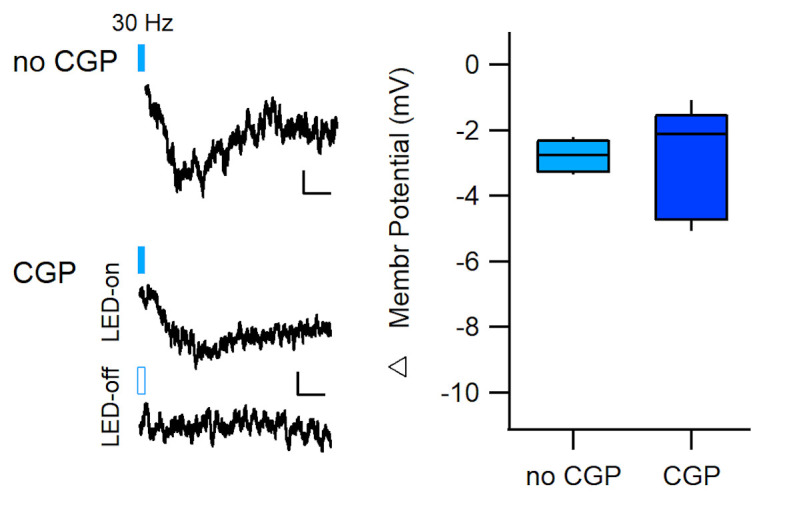
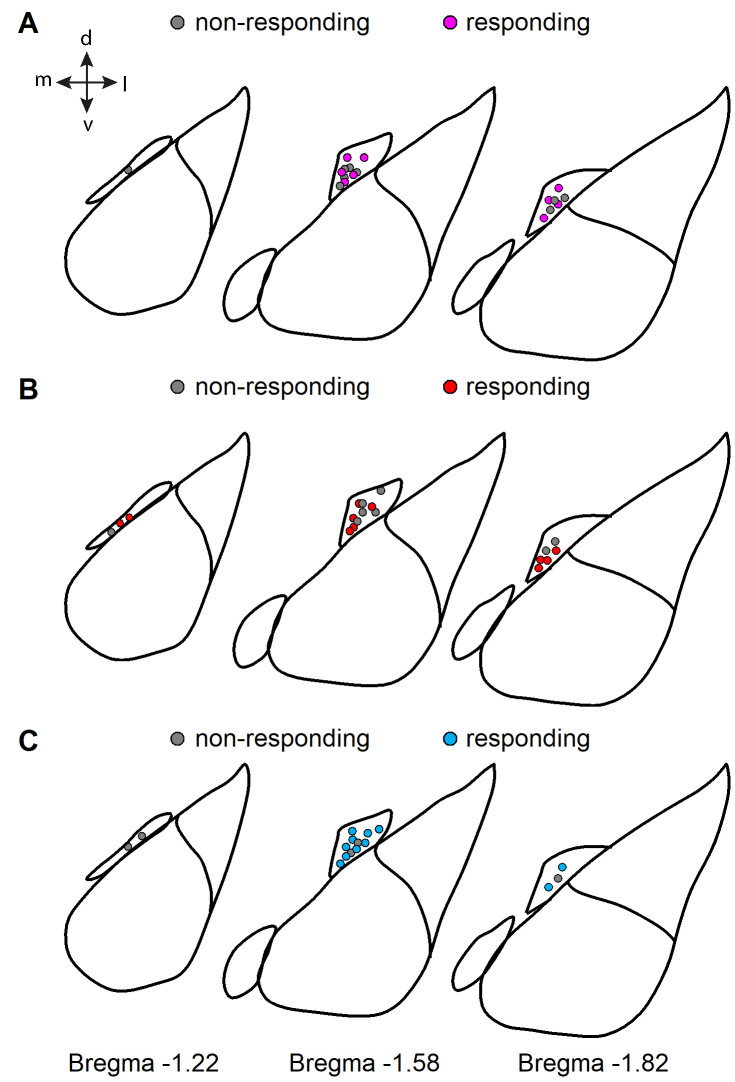
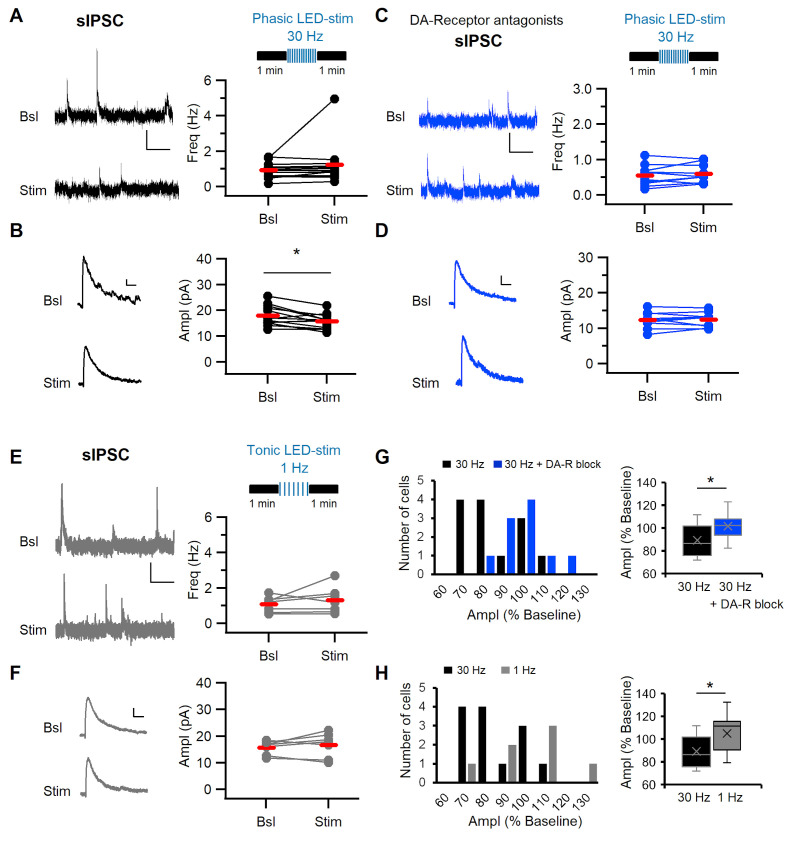
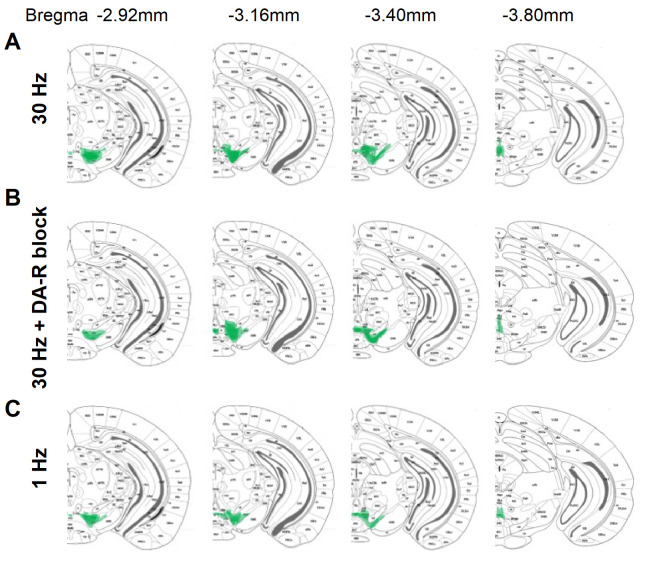
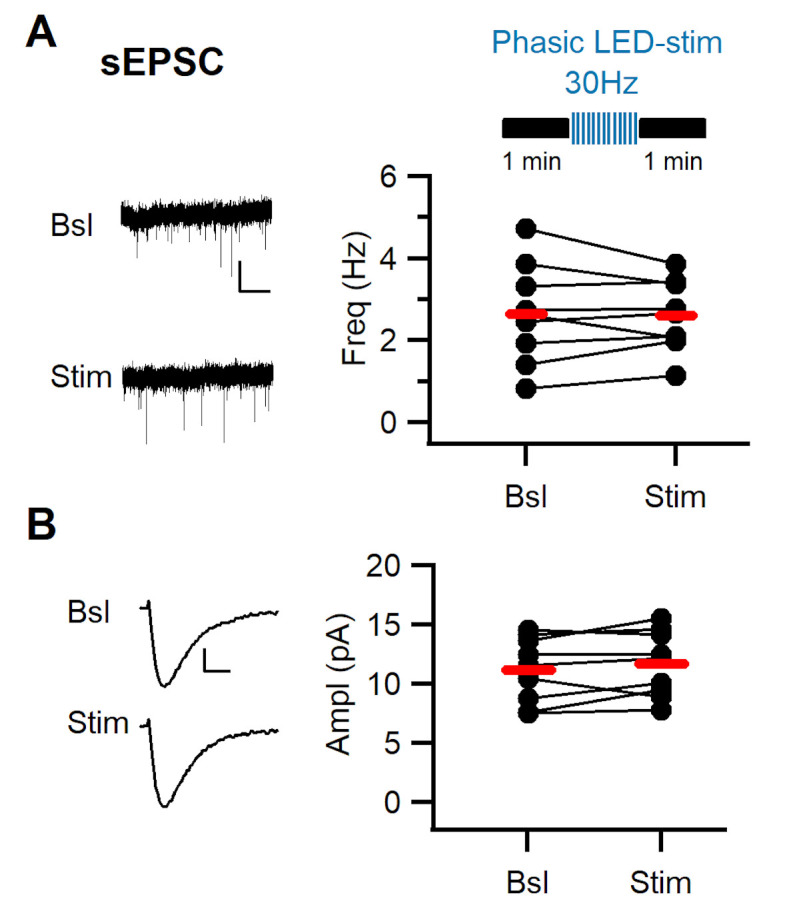
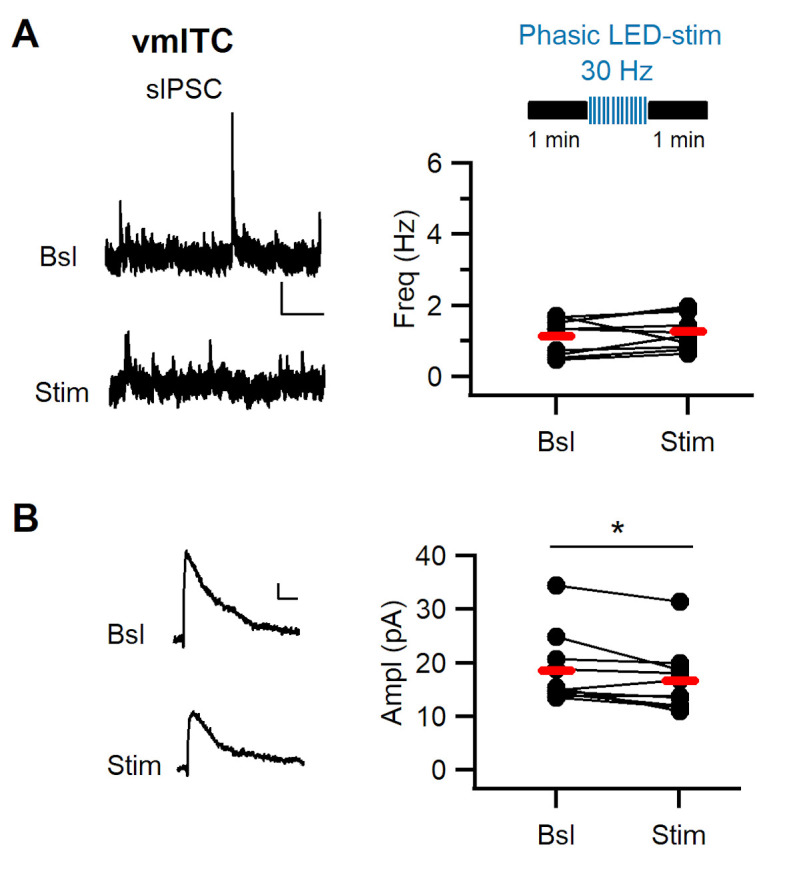
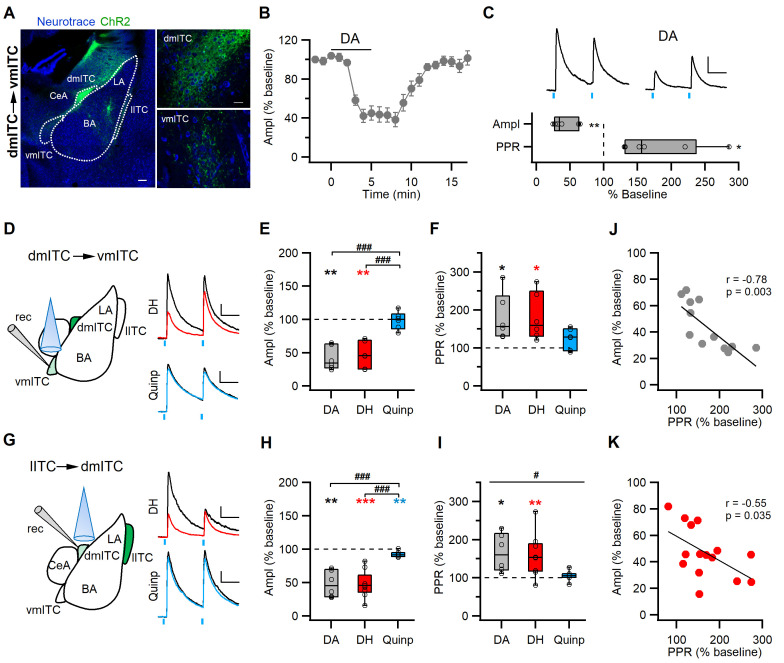
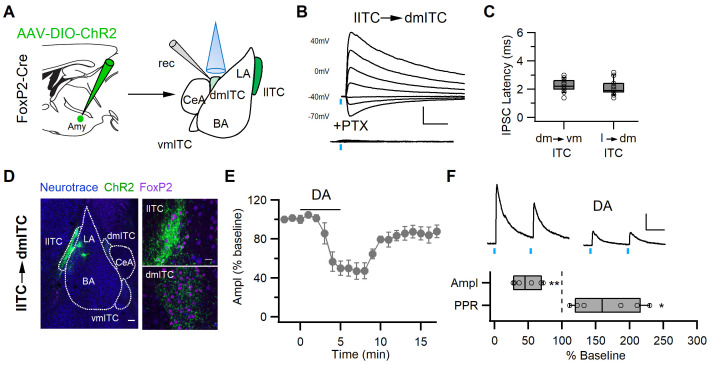
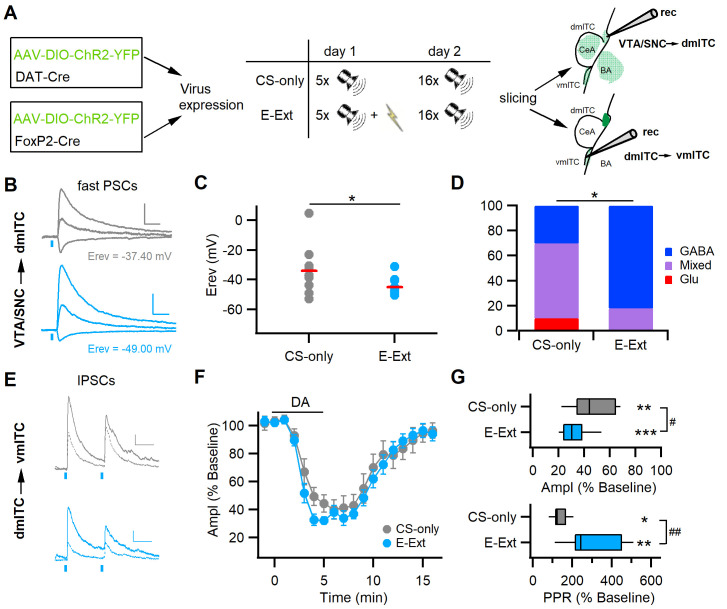
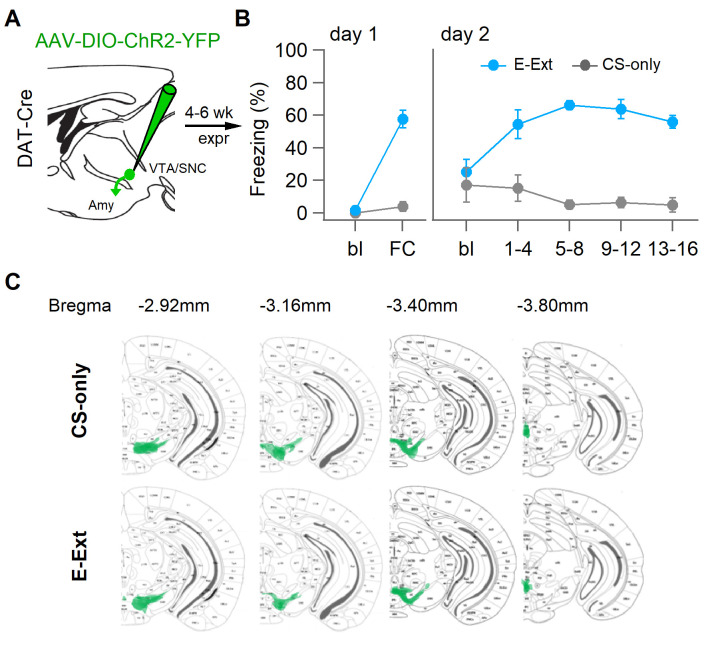
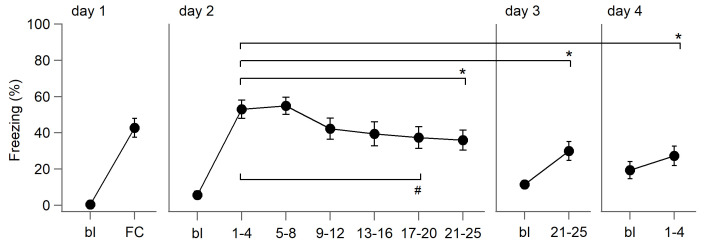
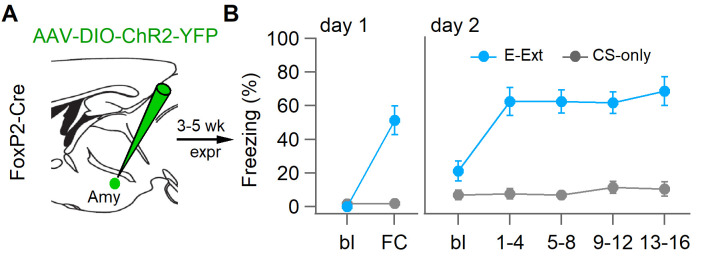
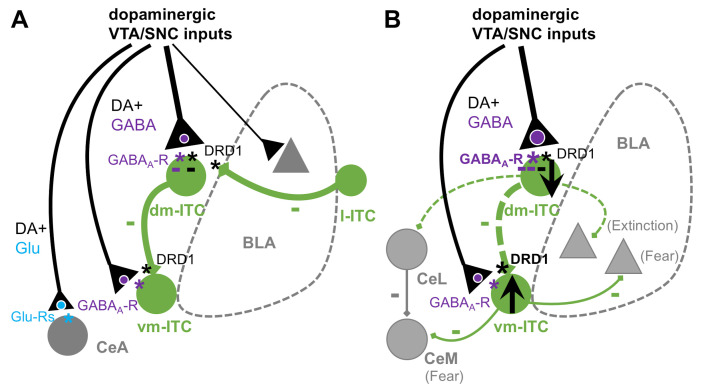
Dopaminergic signaling plays an important role in associative learning, including fear and extinction learning. Dopaminergic midbrain neurons encode prediction error-like signals when threats differ from expectations. Within the amygdala, GABAergic intercalated cell (ITC) clusters receive one of the densest dopaminergic projections, but their physiological consequences are incompletely understood. ITCs are important for fear extinction, a function thought to be supported by activation of ventromedial ITCs that inhibit central amygdala fear output. In mice, we reveal two distinct novel mechanisms by which mesencephalic dopaminergic afferents control ITCs. Firstly, they co-release GABA to mediate rapid, direct inhibition. Secondly, dopamine suppresses inhibitory interactions between distinct ITC clusters via presynaptic D1 receptors. Early extinction training augments both GABA co-release onto dorsomedial ITCs and dopamine-mediated suppression of dorso- to ventromedial inhibition between ITC clusters. These findings provide novel insights into dopaminergic mechanisms shaping the activity balance between distinct ITC clusters that could support their opposing roles in fear behavior.
Keywords: GABA; amygdala; corelease; dopamine; extinction; intercalated neurons; mouse; neuroscience.
© 2021, Aksoy-Aksel et al.
Conflict of interest statement
AA, AG, AS, FF, IE No competing interests declared
Figures
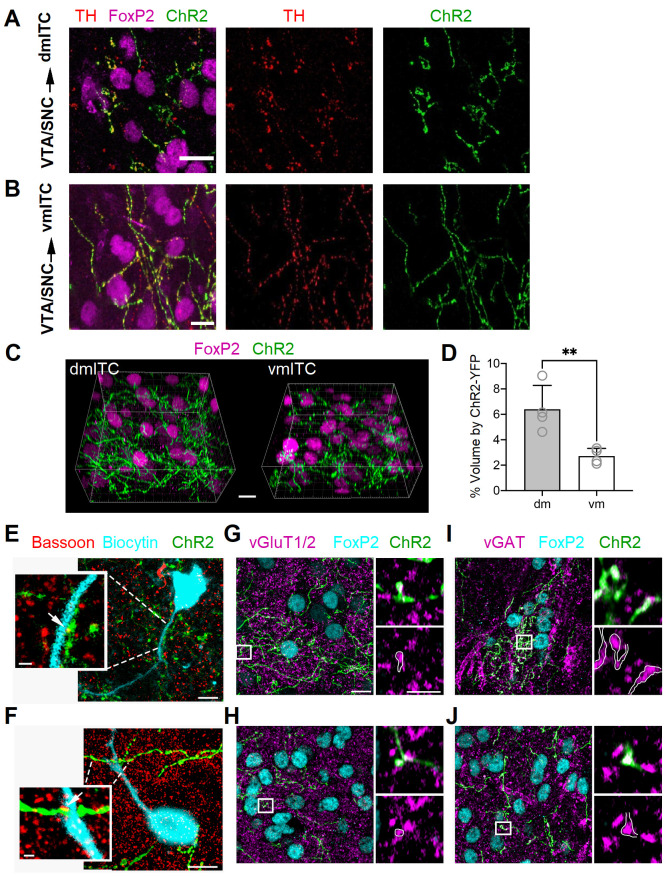
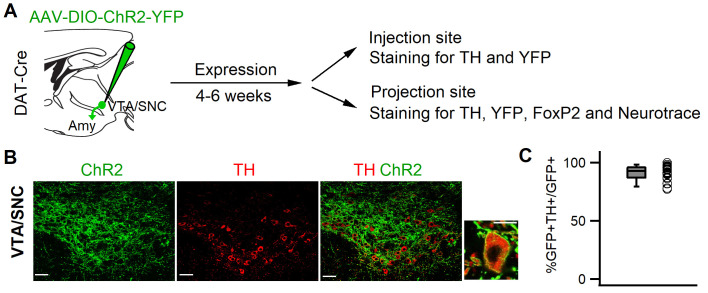
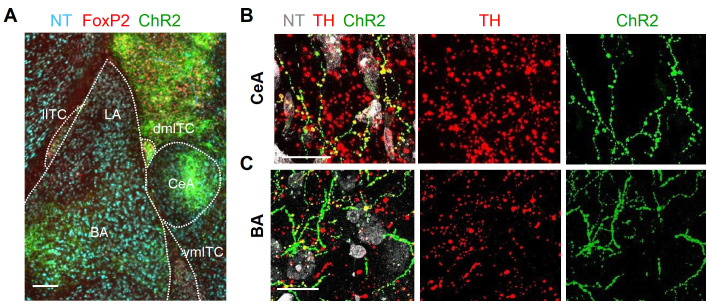
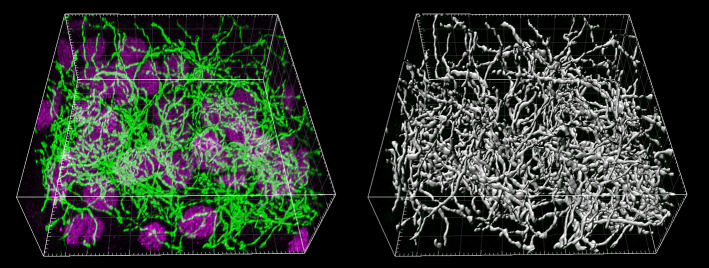
References
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
