

Ligand-dependent kinase activity of MERTK drives efferocytosis in human iPSC-derived macrophages
- PMID: 34035216
- PMCID: PMC8149813
- DOI: 10.1038/s41419-021-03770-0
Ligand-dependent kinase activity of MERTK drives efferocytosis in human iPSC-derived macrophages
Abstract
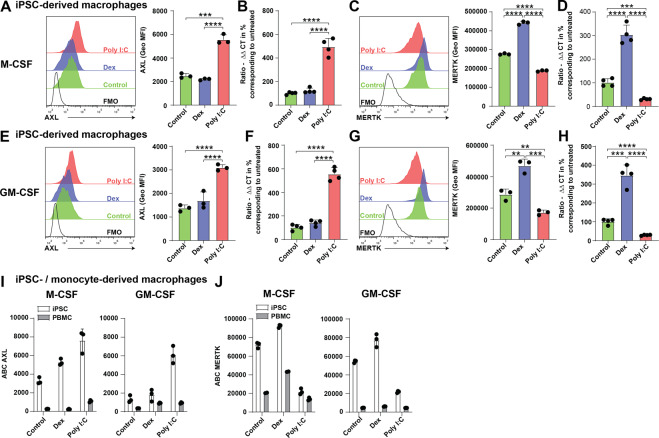
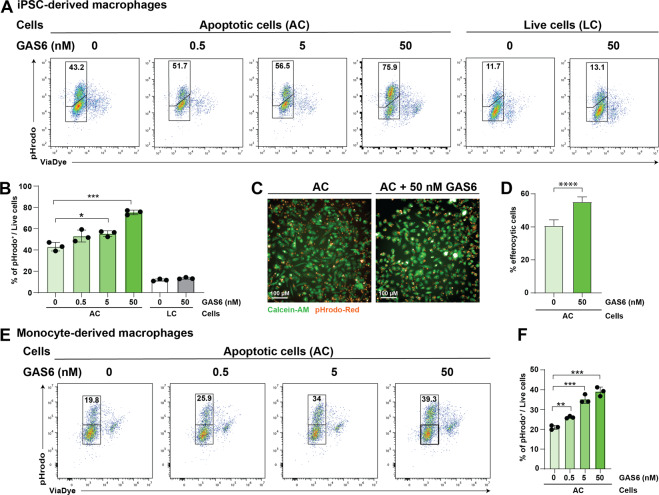
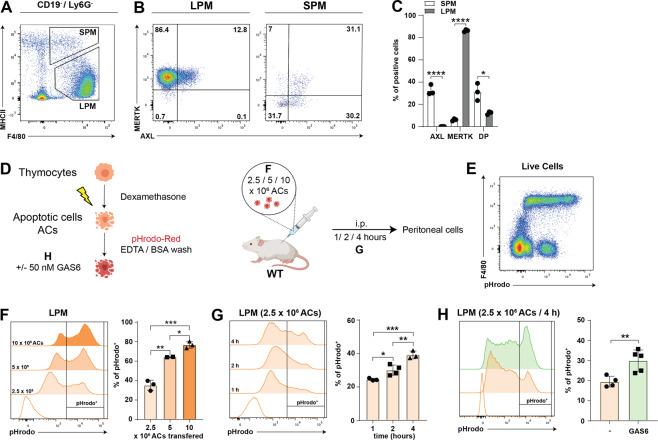
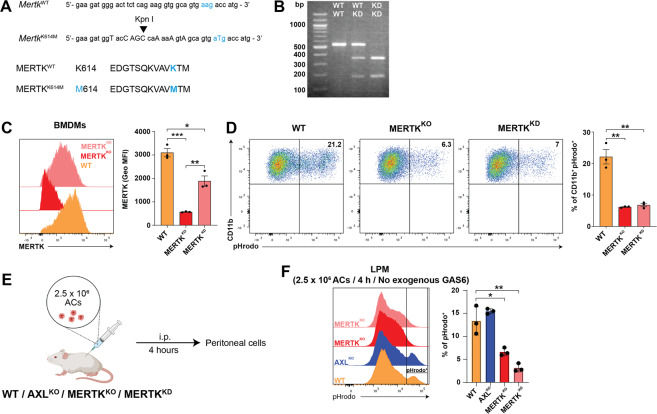
Removal of apoptotic cells by phagocytes (also called efferocytosis) is a crucial process for tissue homeostasis. Professional phagocytes express a plethora of surface receptors enabling them to sense and engulf apoptotic cells, thus avoiding persistence of dead cells and cellular debris and their consequent effects. Dysregulation of efferocytosis is thought to lead to secondary necrosis and associated inflammation and immune activation. Efferocytosis in primarily murine macrophages and dendritic cells has been shown to require TAM RTKs, with MERTK and AXL being critical for clearance of apoptotic cells. The functional role of human orthologs, especially the exact contribution of each individual receptor is less well studied. Here we show that human macrophages differentiated in vitro from iPSC-derived precursor cells express both AXL and MERTK and engulf apoptotic cells. TAM RTK agonism by the natural ligand growth-arrest specific 6 (GAS6) significantly enhanced such efferocytosis. Using a newly-developed mouse model of kinase-dead MERTK, we demonstrate that MERTK kinase activity is essential for efferocytosis in peritoneal macrophages in vivo. Moreover, human iPSC-derived macrophages treated in vitro with blocking antibodies or small molecule inhibitors recapitulated this observation. Hence, our results highlight a conserved MERTK function between mice and humans, and the critical role of its kinase activity in homeostatic efferocytosis.
Conflict of interest statement
The authors whose names are listed in the manuscript certify that they have NO affiliations with or involvement in any organization or entity with any financial interest (such as honoraria; educational grants; participation in speakers’ bureaus; membership, employment, consultancies, stock ownership, or other equity interest; and expert testimony or patent-licensing arrangements), or non-financial interest (such as personal or professional relationships, affiliations, knowledge or beliefs) in the subject matter or materials discussed in this manuscript.
Figures
Similar articles
-
MERTK tyrosine kinase receptor together with TIM4 phosphatidylserine receptor mediates distinct signal transduction pathways for efferocytosis and cell proliferation.J Biol Chem. 2019 May 3;294(18):7221-7230. doi: 10.1074/jbc.RA118.006628. Epub 2019 Mar 7. J Biol Chem. 2019. PMID: 30846565 Free PMC article.
-
Hydroxychloroquine enhances efferocytosis and modulates inflammation via MerTK/Gas6 signaling in a pristane-induced lupus mouse model.Front Immunol. 2025 Jun 16;16:1524315. doi: 10.3389/fimmu.2025.1524315. eCollection 2025. Front Immunol. 2025. PMID: 40589760 Free PMC article.
-
Axl and MerTK receptor tyrosine kinases maintain human macrophage efferocytic capacity in the presence of viral triggers.Eur J Immunol. 2018 May;48(5):855-860. doi: 10.1002/eji.201747283. Epub 2018 Feb 22. Eur J Immunol. 2018. PMID: 29400409 Free PMC article.
-
Tyro3, Axl, Mertk receptor-mediated efferocytosis and immune regulation in the tumor environment.Int Rev Cell Mol Biol. 2021;361:165-210. doi: 10.1016/bs.ircmb.2021.02.002. Epub 2021 May 10. Int Rev Cell Mol Biol. 2021. PMID: 34074493 Review.
-
The role of TAM family receptors and ligands in the nervous system: From development to pathobiology.Pharmacol Ther. 2018 Aug;188:97-117. doi: 10.1016/j.pharmthera.2018.03.002. Epub 2018 Mar 4. Pharmacol Ther. 2018. PMID: 29514053 Free PMC article. Review.
Cited by
-
Elevated expression of macrophage MERTK exhibits profibrotic effects and results in defective regulation of efferocytosis function in pulmonary fibrosis.Respir Res. 2023 Apr 29;24(1):118. doi: 10.1186/s12931-023-02424-3. Respir Res. 2023. PMID: 37120511 Free PMC article.
-
ACK1 and BRK non-receptor tyrosine kinase deficiencies are associated with familial systemic lupus and involved in efferocytosis.medRxiv [Preprint]. 2024 Jun 5:2024.02.15.24302255. doi: 10.1101/2024.02.15.24302255. medRxiv. 2024. Update in: Elife. 2024 Nov 21;13:RP96085. doi: 10.7554/eLife.96085. PMID: 38883731 Free PMC article. Updated. Preprint.
-
Gas6 induces AIM to suppress acute lung injury in mice by inhibiting NLRP3 inflammasome activation and inducing autophagy.Front Immunol. 2025 Feb 17;16:1523166. doi: 10.3389/fimmu.2025.1523166. eCollection 2025. Front Immunol. 2025. PMID: 40034700 Free PMC article.
-
Efferocytosis by Macrophages Attenuates Inflammatory Responses Following Ultraviolet B-Induced Apoptosis in Corneal Stromal Cells.Invest Ophthalmol Vis Sci. 2025 Jul 1;66(9):49. doi: 10.1167/iovs.66.9.49. Invest Ophthalmol Vis Sci. 2025. PMID: 40673741 Free PMC article.
-
Enhancing therapeutic anti-cancer responses by combining immune checkpoint and tyrosine kinase inhibition.Mol Cancer. 2022 Sep 29;21(1):189. doi: 10.1186/s12943-022-01656-z. Mol Cancer. 2022. PMID: 36175961 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
