

Caerulomycin and collismycin antibiotics share a trans-acting flavoprotein-dependent assembly line for 2,2'-bipyridine formation
- PMID: 34035275
- PMCID: PMC8149447
- DOI: 10.1038/s41467-021-23475-4
Caerulomycin and collismycin antibiotics share a trans-acting flavoprotein-dependent assembly line for 2,2'-bipyridine formation
Abstract
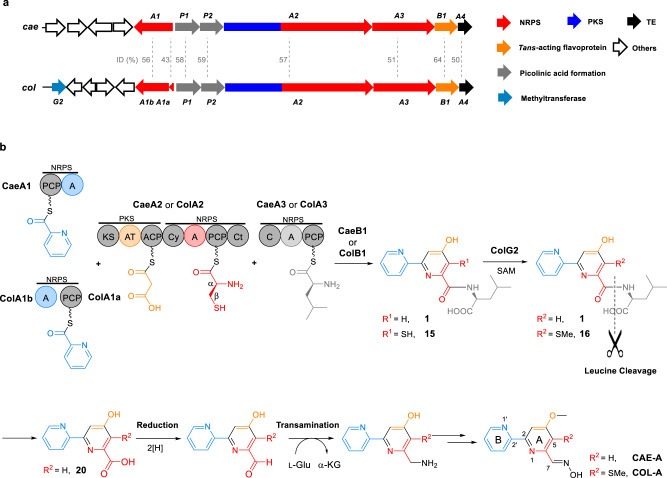
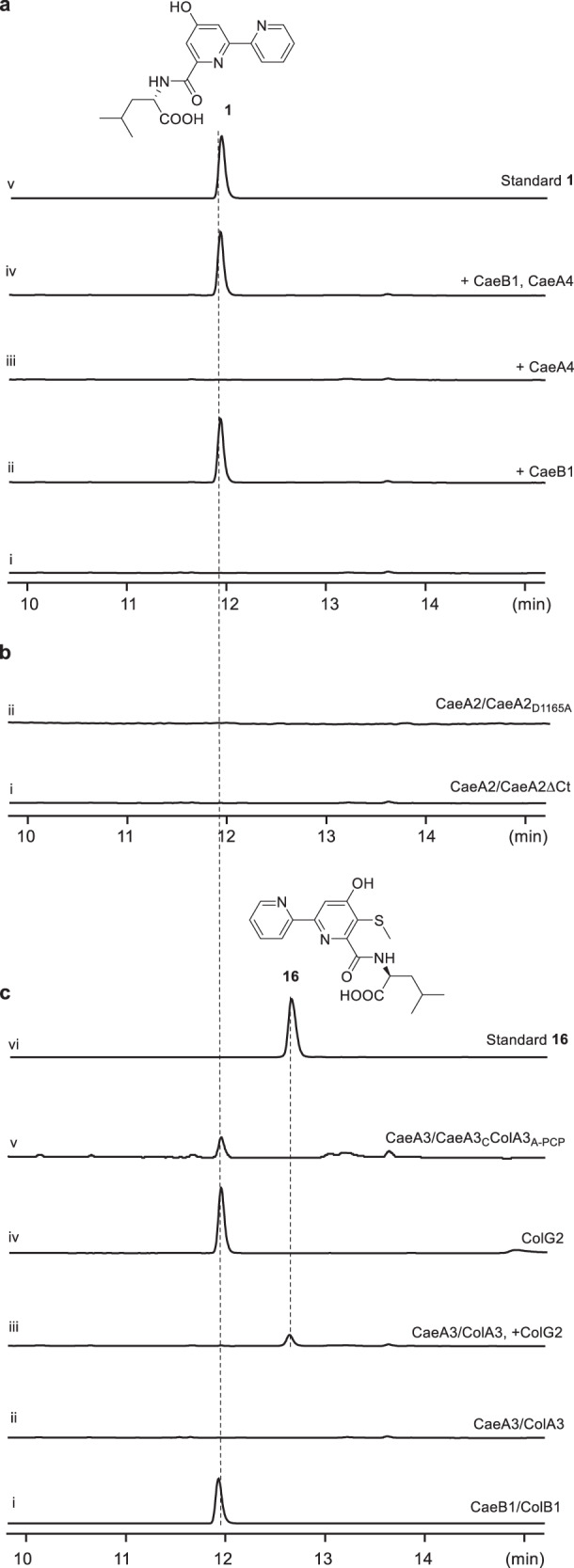
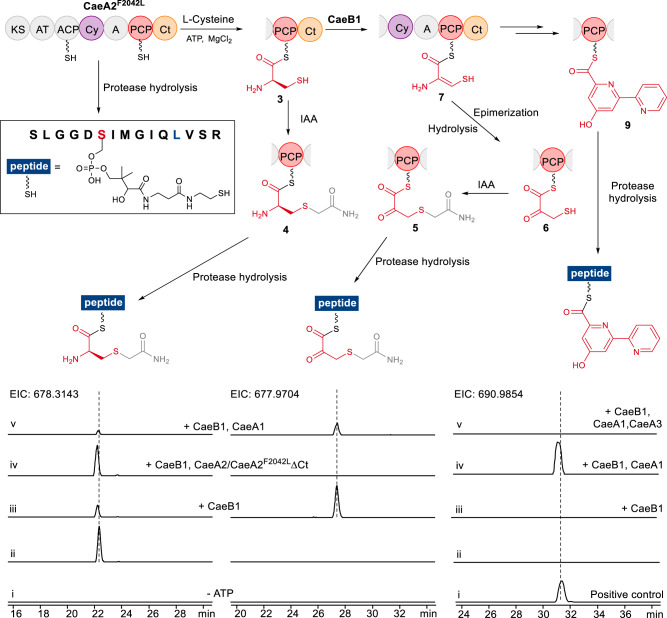
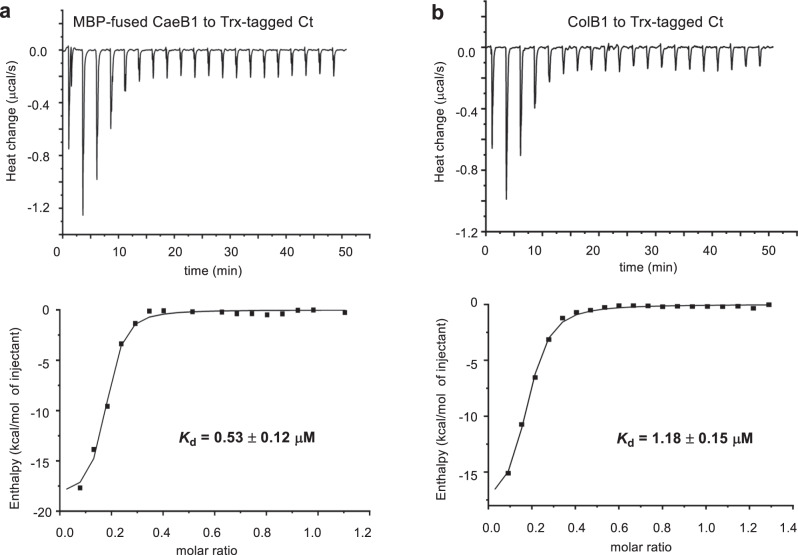
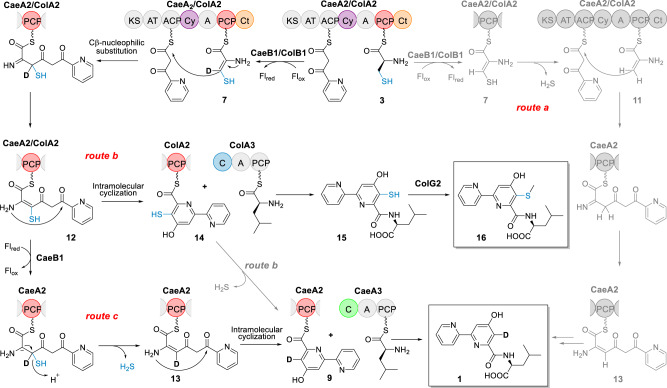
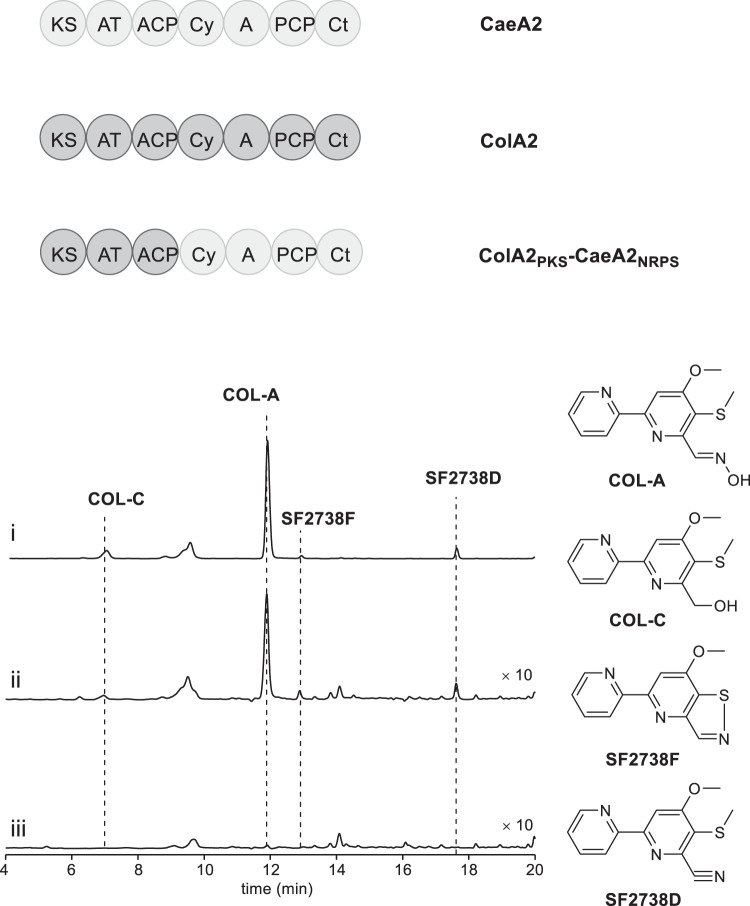
Linear nonribosomal peptide synthetases (NRPSs) and polyketide synthases (PKSs) template the modular biosynthesis of numerous nonribosomal peptides, polyketides and their hybrids through assembly line chemistry. This chemistry can be complex and highly varied, and thus challenges our understanding in NRPS and PKS-programmed, diverse biosynthetic processes using amino acid and carboxylate building blocks. Here, we report that caerulomycin and collismycin peptide-polyketide hybrid antibiotics share an assembly line that involves unusual NRPS activity to engage a trans-acting flavoprotein in C-C bond formation and heterocyclization during 2,2'-bipyridine formation. Simultaneously, this assembly line provides dethiolated and thiolated 2,2'-bipyridine intermediates through differential treatment of the sulfhydryl group arising from L-cysteine incorporation. Subsequent L-leucine extension, which does not contribute any atoms to either caerulomycins or collismycins, plays a key role in sulfur fate determination by selectively advancing one of the two 2,2'-bipyridine intermediates down a path to the final products with or without sulfur decoration. These findings further the appreciation of assembly line chemistry and will facilitate the development of related molecules using synthetic biology approaches.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Caerulomycins and collismycins share a common paradigm for 2,2'-bipyridine biosynthesis via an unusual hybrid polyketide-peptide assembly Logic.J Am Chem Soc. 2012 Jun 6;134(22):9038-41. doi: 10.1021/ja3016457. Epub 2012 May 23. J Am Chem Soc. 2012. PMID: 22594451
-
Elucidating the biosynthetic pathway for the polyketide-nonribosomal peptide collismycin A: mechanism for formation of the 2,2'-bipyridyl ring.Chem Biol. 2012 Mar 23;19(3):399-413. doi: 10.1016/j.chembiol.2012.01.014. Chem Biol. 2012. PMID: 22444595
-
Domain-Targeted Metabolomics Delineates the Heterocycle Assembly Steps of Colibactin Biosynthesis.J Am Chem Soc. 2017 Mar 22;139(11):4195-4201. doi: 10.1021/jacs.7b00659. Epub 2017 Mar 10. J Am Chem Soc. 2017. PMID: 28240912 Free PMC article.
-
Protein-protein interactions in polyketide synthase-nonribosomal peptide synthetase hybrid assembly lines.Nat Prod Rep. 2018 Nov 14;35(11):1185-1209. doi: 10.1039/c8np00022k. Nat Prod Rep. 2018. PMID: 30074030 Review.
-
Discovery of caerulomycin/collismycin-type 2,2'-bipyridine natural products in the genomic era.J Ind Microbiol Biotechnol. 2019 Mar;46(3-4):459-468. doi: 10.1007/s10295-018-2092-7. Epub 2018 Nov 27. J Ind Microbiol Biotechnol. 2019. PMID: 30484122 Review.
Cited by
-
AI-assisted isolation of bioactive Dipyrimicins from Amycolatopsis azurea and identification of its corresponding dip biosynthetic gene cluster.bioRxiv [Preprint]. 2025 Mar 21:2025.03.21.644653. doi: 10.1101/2025.03.21.644653. bioRxiv. 2025. PMID: 40166164 Free PMC article. Preprint.
-
Advances on the biosynthesis of pyridine rings.Eng Microbiol. 2022 Dec 13;3(2):100064. doi: 10.1016/j.engmic.2022.100064. eCollection 2023 Jun. Eng Microbiol. 2022. PMID: 39629243 Free PMC article. Review.
-
Investigation of Optical Properties and Biological Activities of Antimony(III) Halide Templated by 2,2'-Bipyridinium for Optoelectronic Devices.ACS Omega. 2025 Apr 29;10(18):19107-19114. doi: 10.1021/acsomega.5c01763. eCollection 2025 May 13. ACS Omega. 2025. PMID: 40385195 Free PMC article.
-
Comprehensive safety and toxicity analysis of 2,2'-Bipyridine derivatives in combating MRSA biofilm formation and persistence.Front Cell Infect Microbiol. 2025 Jan 24;15:1493679. doi: 10.3389/fcimb.2025.1493679. eCollection 2025. Front Cell Infect Microbiol. 2025. PMID: 39925377 Free PMC article.
-
Discovery and Characterization of Oxazolismycin, an Oxazole-Containing Natural Product with an Atypical Loading Mechanism.JACS Au. 2025 Jun 17;5(7):3000-3007. doi: 10.1021/jacsau.5c00326. eCollection 2025 Jul 28. JACS Au. 2025. PMID: 40747025 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
