

Pleiotropic Effect of Hormone Insulin-Like Growth Factor-I in Immune Response and Pathogenesis in Leishmaniases
- PMID: 34036108
- PMCID: PMC8116165
- DOI: 10.1155/2021/6614475
Pleiotropic Effect of Hormone Insulin-Like Growth Factor-I in Immune Response and Pathogenesis in Leishmaniases
Abstract
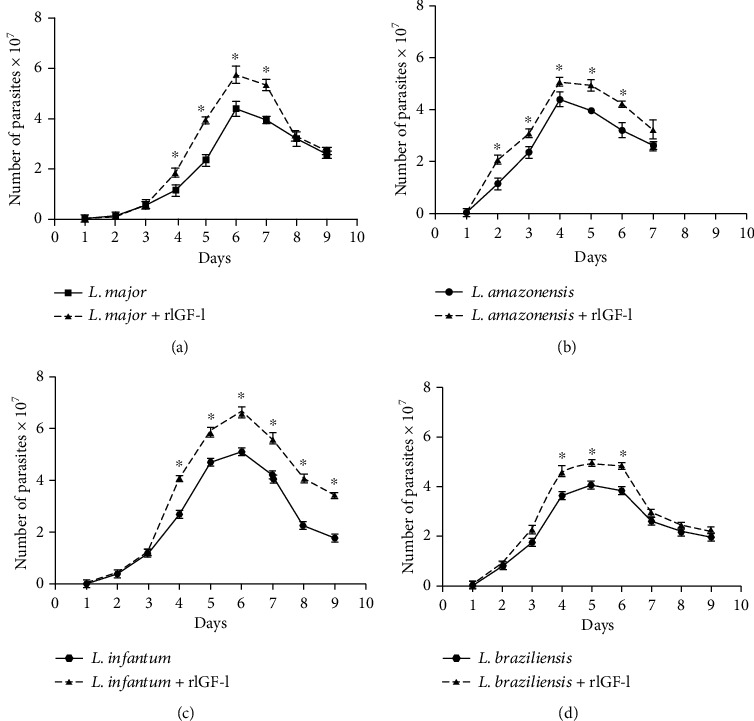
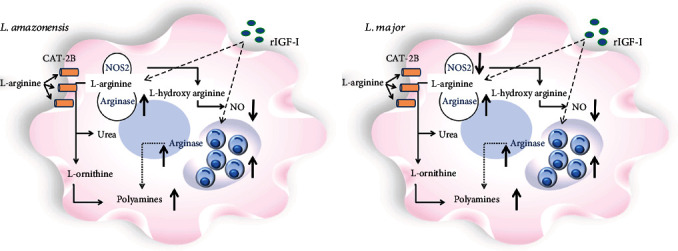
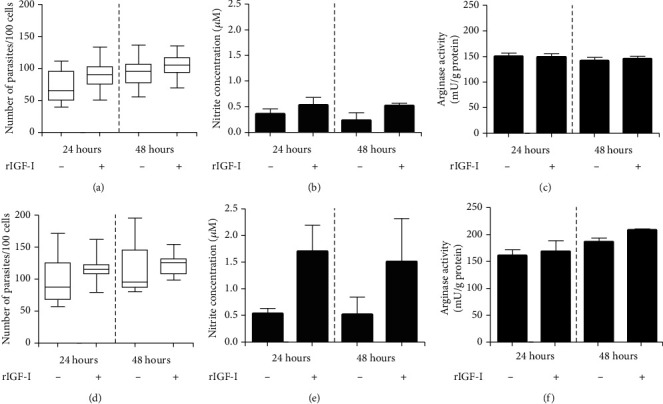
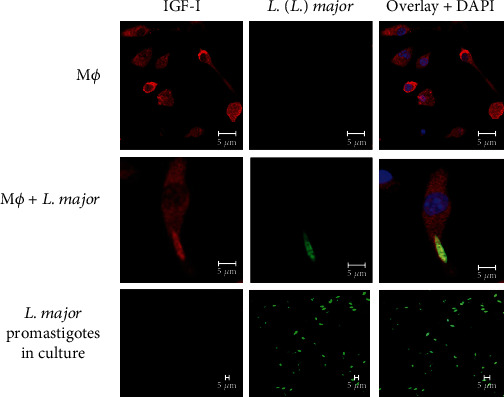
Leishmaniases are diseases caused by several Leishmania species, and many factors contribute to the development of the infection. Because the adaptive immune response does not fully explain the outcome of Leishmania infection and considering that the initial events are crucial in the establishment of the infection, we investigated one of the growth factors, the insulin-like growth factor-I (IGF-I), found in circulation and produced by different cells including macrophages and present in the skin where the parasite is inoculated. Here, we review the role of IGF-I in leishmaniasis experimental models and human patients. IGF-I induces the growth of different Leishmania species in vitro and alters the disease outcome increasing the parasite load and lesion size, especially in L. major- and L. amazonensis-infected mouse leishmaniasis. IGF-I affects the parasite interacting with the IGF-I receptor present on Leishmania. During Leishmania-macrophage interaction, IGF-I acts on the arginine metabolic pathway, resulting in polyamine production both in macrophages and Leishmania. IGF-I and cytokines interact with reciprocal influences on their expression. IL-4 is a hallmark of susceptibility to L. major in murine leishmaniasis, but we observed that IGF-I operates astoundingly as an effector element of the IL-4. Approaching human leishmaniasis, patients with mucosal, disseminated, and visceral diseases presented surprisingly low IGF-I serum levels, suggesting diverse effects than parasite growth. We observed that low IGF-I levels might contribute to the inflammatory response persistence and delayed lesion healing in human cutaneous leishmaniasis and the anemia development in visceral leishmaniasis. We must highlight the complexity of infection revealed depending on the Leishmania species and the parasite's developmental stages. Because IGF-I exerts pleiotropic effects on the biology of interaction and disease pathogenesis, IGF-I turns up as an attractive tool to explore biological and pathogenic processes underlying infection development. IGF-I pleiotropic effects open further the possibility of approaching IGF-I as a therapeutical target.
Copyright © 2021 Luiza C. Reis et al.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
Similar articles
-
Dual Role of Insulin-Like Growth Factor (IGF)-I in American Tegumentary Leishmaniasis.J Immunol Res. 2021 Mar 29;2021:6657785. doi: 10.1155/2021/6657785. eCollection 2021. J Immunol Res. 2021. PMID: 33860062 Free PMC article.
-
Insulin-like growth factor-I serum levels and their biological effects on Leishmania isolates from different clinical forms of American tegumentary leishmaniasis.Parasit Vectors. 2016 Jun 11;9(1):335. doi: 10.1186/s13071-016-1619-x. Parasit Vectors. 2016. PMID: 27286813 Free PMC article.
-
Effect of insulin-like growth factor-I on Leishmania amazonensis promastigote arginase activation and reciprocal inhibition of NOS2 pathway in macrophage in vitro.Scand J Immunol. 2007 Aug-Sep;66(2-3):287-96. doi: 10.1111/j.1365-3083.2007.01950.x. Scand J Immunol. 2007. PMID: 17635806
-
Interaction of Leishmania with the host macrophage.Trends Parasitol. 2002 Aug;18(8):332-4. doi: 10.1016/s1471-4922(02)02352-8. Trends Parasitol. 2002. PMID: 12377273 Review.
-
Leishmania species-dependent functional duality of toll-like receptor 2.IUBMB Life. 2019 Nov;71(11):1685-1700. doi: 10.1002/iub.2129. Epub 2019 Jul 22. IUBMB Life. 2019. PMID: 31329370 Review.
Cited by
-
miR-548d-3p Is Up-Regulated in Human Visceral Leishmaniasis and Suppresses Parasite Growth in Macrophages.Front Cell Infect Microbiol. 2022 Feb 10;12:826039. doi: 10.3389/fcimb.2022.826039. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 35265535 Free PMC article.
-
miR-548d-3p Alters Parasite Growth and Inflammation in Leishmania (Viannia) braziliensis Infection.Front Cell Infect Microbiol. 2021 Jun 10;11:687647. doi: 10.3389/fcimb.2021.687647. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 34178725 Free PMC article.
References
-
- Gumy A., Louis J. A., Launois P. The murine model of infection with Leishmania major and its importance for the deciphering of mechanisms underlying differences in Th cell differentiation in mice from different genetic backgrounds. International Journal for Parasitology. 2004;34(4):433–444. doi: 10.1016/j.ijpara.2003.11.021. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
