

Regulation of aquaporin-4 expression in the central nervous system investigated using M23-AQP4 null mouse
- PMID: 34038017
- PMCID: PMC8361696
- DOI: 10.1002/glia.24032
Regulation of aquaporin-4 expression in the central nervous system investigated using M23-AQP4 null mouse
Abstract
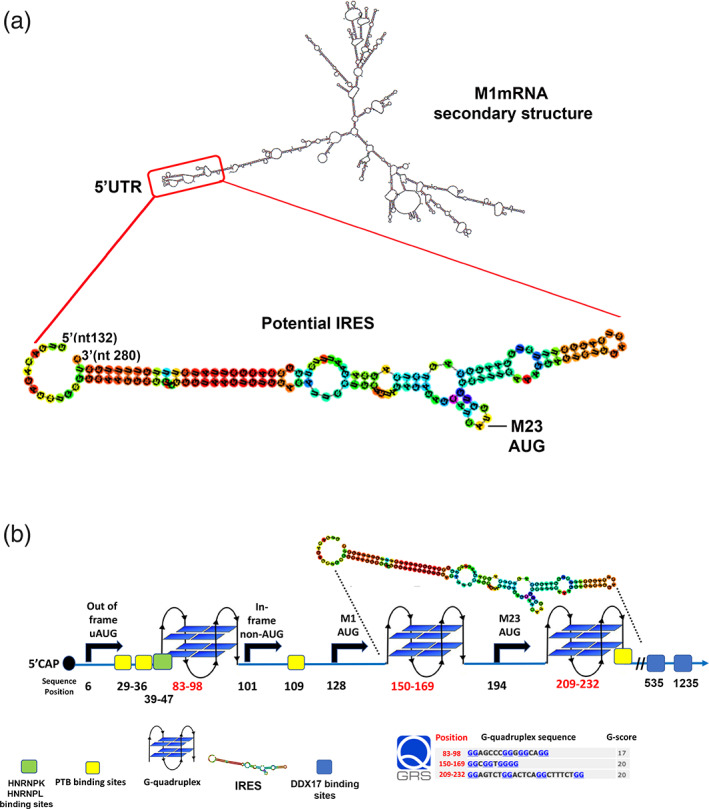
In astrocytes, unknown mechanisms regulate the expression of M1 and M23 isoforms of water channel aquaporin-4 (M1-AQP4 and M23-AQP4). The ratio between these two isoforms controls the AQP4 assembly state in the plasma membrane known as orthogonal arrays of particles (OAPs). To give new insights into these mechanisms, here, we explore the regulation of AQP4 expression in the spinal cord of a CRISPR/Cas9 M23-null mouse model (M23-null). In the M23-null spinal cord OAP assembly, the perivascular localization of AQP4 and M1-AQP4 protein were drastically reduced. In heterozygous, M1-AQP4 was proportionally reduced with M23-AQP4, maintaining the isoform ratio unaffected. We hypothesize a role of the M23-AQP4 in the regulation of M1-AQP4 expression. M1-AQP4 transcription, splicing and M1-AQP4 protein degradation were found to be unaffected in M23-null spinal cord and in M23-null astrocyte primary culture. The translational control was investigated by mRNA-protein pull down and quantitative mass spectrometry, to isolate and quantify AQP4 mRNA binding proteins (AQP4-RBPs). Compared to WT, in M23-null spinal cord, the interaction between AQP4 mRNA and polypyrimidine tract binding protein 1, a positive regulator of AQP4 translation, was higher, while interaction with the RNA helicase DDX17 was lower. In astrocyte primary cultures, DDX17 knockdown upregulated AQP4 protein expression and increased cell swelling, leaving AQP4 mRNA levels unchanged. Here, we identify AQP4-RBPs and provide evidence that in mouse spinal cord M23-AQP4 deletion changes the interaction between AQP4 mRNA and some RBPs involved in AQP4 translation. We describe for the first time the RNA helicase DDX17 as a regulator of AQP4 expression in astrocytes.
Keywords: AQP4 mRNA binding proteins; CRISPR/Cas9 M23-null mouse model; DDX17; PTBP1; translational regulation.
© 2021 The Authors. GLIA published by Wiley Periodicals LLC.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
-
- Abe, Y., Ikeshima‐Kataoka, H., Goda, W., Niikura, T., & Yasui, M. (2012). An astrocyte‐specific enhancer of the aquaporin‐4 gene functions through a consensus sequence of POU transcription factors in concert with multiple upstream elements. Journal of Neurochemistry, 120(6), 899–912. 10.1111/j.1471-4159.2012.07652.x - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
