

Chemokines act as phosphatidylserine-bound "find-me" signals in apoptotic cell clearance
- PMID: 34038417
- PMCID: PMC8213124
- DOI: 10.1371/journal.pbio.3001259
Chemokines act as phosphatidylserine-bound "find-me" signals in apoptotic cell clearance
Abstract
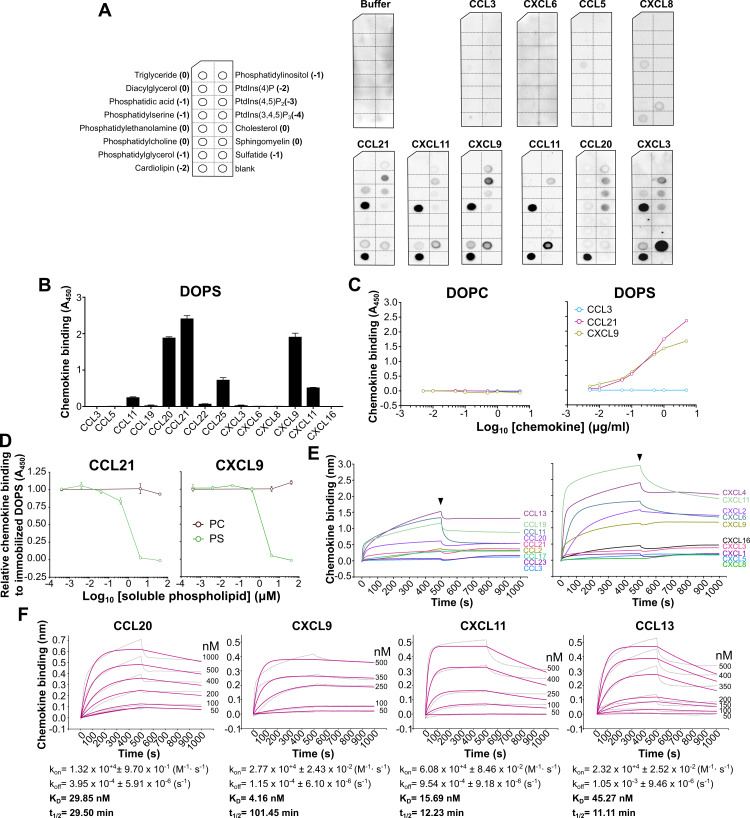
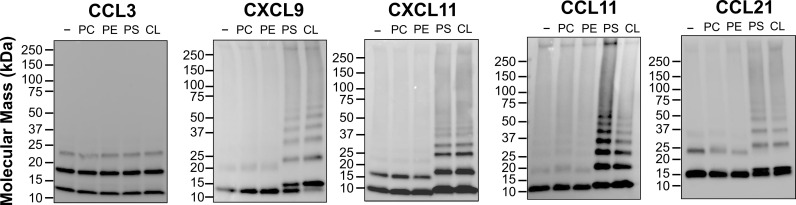
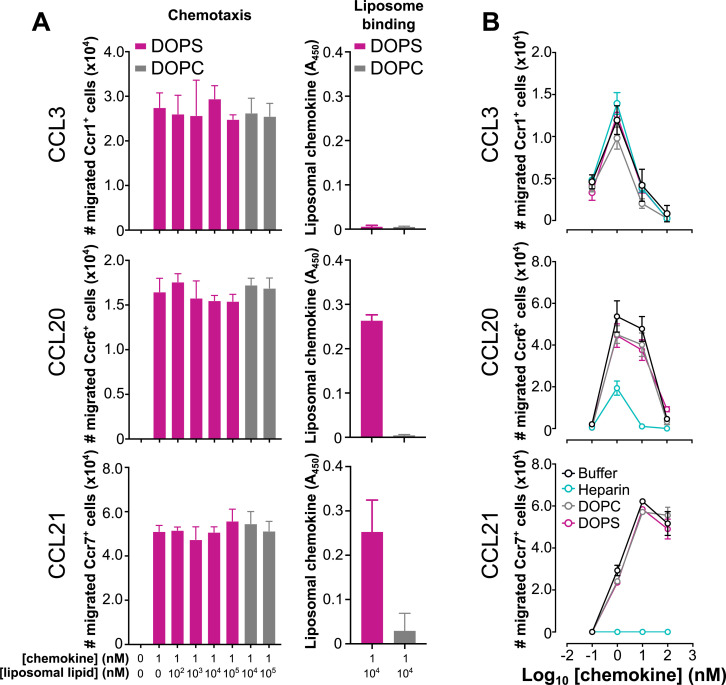
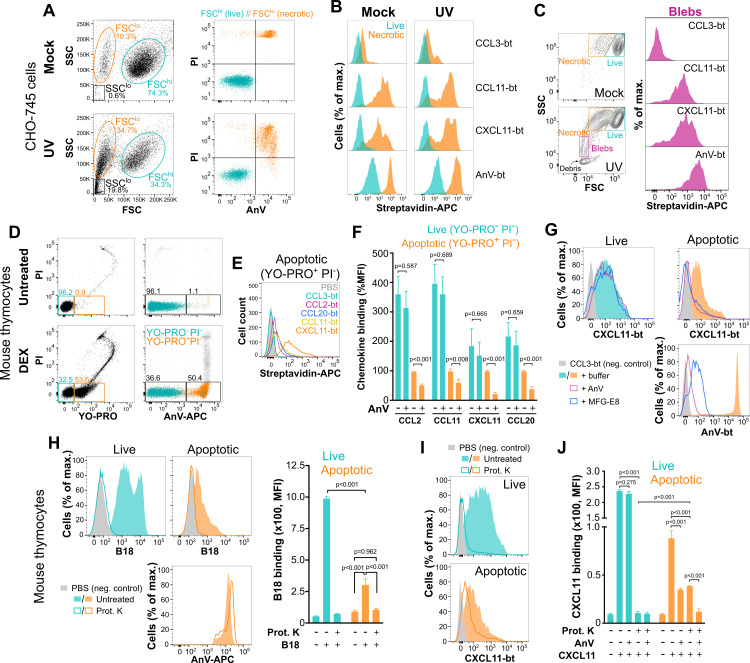
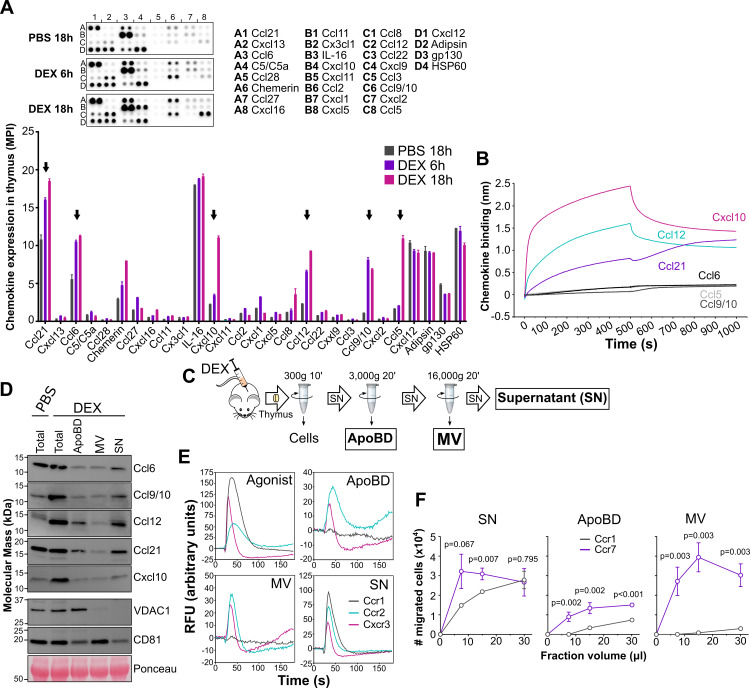
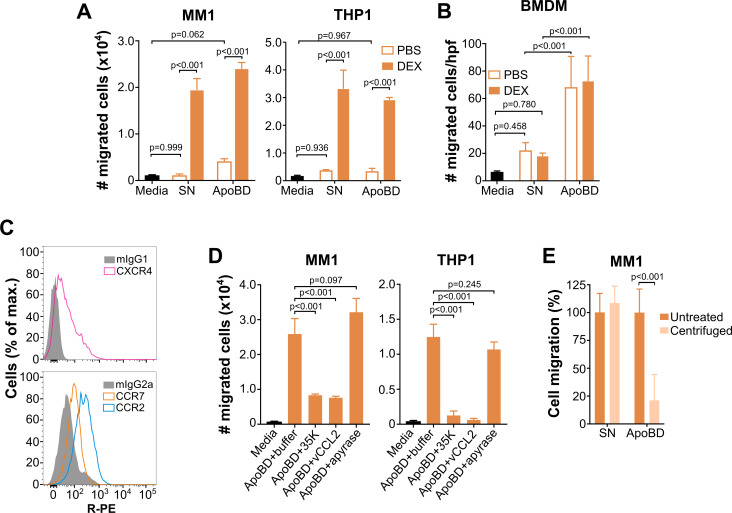
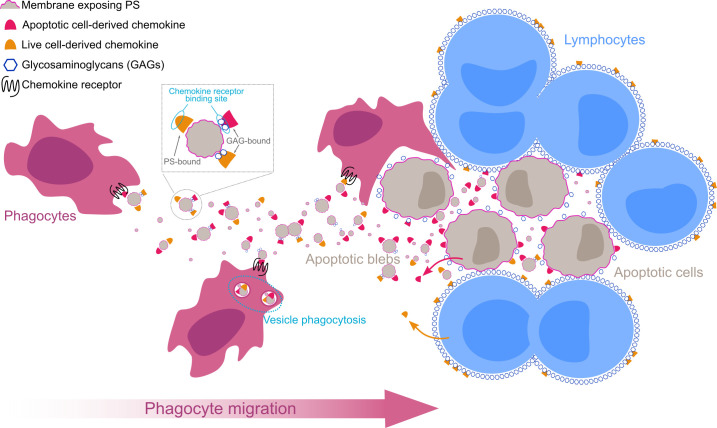
Removal of apoptotic cells is essential for maintenance of tissue homeostasis. Chemotactic cues termed "find-me" signals attract phagocytes toward apoptotic cells, which selectively expose the anionic phospholipid phosphatidylserine (PS) and other "eat-me" signals to distinguish healthy from apoptotic cells for phagocytosis. Blebs released by apoptotic cells can deliver find-me signals; however, the mechanism is poorly understood. Here, we demonstrate that apoptotic blebs generated in vivo from mouse thymus attract phagocytes using endogenous chemokines bound to the bleb surface. We show that chemokine binding to apoptotic cells is mediated by PS and that high affinity binding of PS and other anionic phospholipids is a general property of many but not all chemokines. Chemokines are positively charged proteins that also bind to anionic glycosaminoglycans (GAGs) on cell surfaces for presentation to leukocyte G protein-coupled receptors (GPCRs). We found that apoptotic cells down-regulate GAGs as they up-regulate PS on the cell surface and that PS-bound chemokines, unlike GAG-bound chemokines, are able to directly activate chemokine receptors. Thus, we conclude that PS-bound chemokines may serve as find-me signals on apoptotic vesicles acting at cognate chemokine receptors on leukocytes.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Surface code--biophysical signals for apoptotic cell clearance.Phys Biol. 2013 Dec;10(6):065007. doi: 10.1088/1478-3975/10/6/065007. Epub 2013 Dec 4. Phys Biol. 2013. PMID: 24305041 Review.
-
Externalized phosphatidylinositides on apoptotic cells are eat-me signals recognized by CD14.Cell Death Differ. 2022 Jul;29(7):1423-1432. doi: 10.1038/s41418-022-00931-2. Epub 2022 Jan 11. Cell Death Differ. 2022. PMID: 35017647 Free PMC article.
-
Nucleotides released by apoptotic cells act as a find-me signal to promote phagocytic clearance.Nature. 2009 Sep 10;461(7261):282-6. doi: 10.1038/nature08296. Nature. 2009. PMID: 19741708 Free PMC article.
-
The role of phosphatidylserine recognition receptors in multiple biological functions.Cell Mol Biol Lett. 2020 Mar 26;25:23. doi: 10.1186/s11658-020-00214-z. eCollection 2020. Cell Mol Biol Lett. 2020. PMID: 32226456 Free PMC article. Review.
-
Plasma membrane alterations during apoptosis: role in corpse clearance.Antioxid Redox Signal. 2004 Apr;6(2):269-75. doi: 10.1089/152308604322899332. Antioxid Redox Signal. 2004. PMID: 15025928 Review.
Cited by
-
Phosphatidylserine-Exposing Annexin A1-Positive Extracellular Vesicles: Potential Cancer Biomarkers.Vaccines (Basel). 2023 Mar 13;11(3):639. doi: 10.3390/vaccines11030639. Vaccines (Basel). 2023. PMID: 36992223 Free PMC article.
-
Extracellular vesicles arising from apoptosis: forms, functions, and applications.J Pathol. 2023 Aug;260(5):592-608. doi: 10.1002/path.6138. Epub 2023 Jun 9. J Pathol. 2023. PMID: 37294158 Free PMC article. Review.
-
Apoptotic MSCs and MSC-Derived Apoptotic Bodies as New Therapeutic Tools.Curr Issues Mol Biol. 2022 Oct 24;44(11):5153-5172. doi: 10.3390/cimb44110351. Curr Issues Mol Biol. 2022. PMID: 36354663 Free PMC article. Review.
-
Events Occurring in the Axotomized Facial Nucleus.Cells. 2022 Jun 29;11(13):2068. doi: 10.3390/cells11132068. Cells. 2022. PMID: 35805151 Free PMC article. Review.
-
ApoBDs: a paradigm shift from cellular debris to therapeutic vehicles.Front Endocrinol (Lausanne). 2025 Jul 17;16:1626796. doi: 10.3389/fendo.2025.1626796. eCollection 2025. Front Endocrinol (Lausanne). 2025. PMID: 40747304 Free PMC article. Review.
References
-
- Bachelerie F, Ben-Baruch A, Burkhardt AM, Combadiere C, Farber JM, et al.. International Union of Basic and Clinical Pharmacology. [corrected]. LXXXIX. Update on the extended family of chemokine receptors and introducing a new nomenclature for atypical chemokine receptors. Pharmacol Rev. 2014;66:1–79. doi: 10.1124/pr.113.007724 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
