

Disruption of paternal circadian rhythm affects metabolic health in male offspring via nongerm cell factors
- PMID: 34039610
- PMCID: PMC8153725
- DOI: 10.1126/sciadv.abg6424
Disruption of paternal circadian rhythm affects metabolic health in male offspring via nongerm cell factors
Abstract
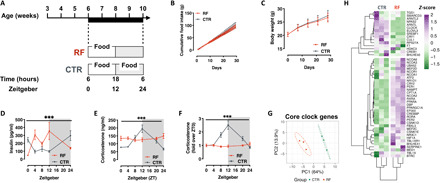
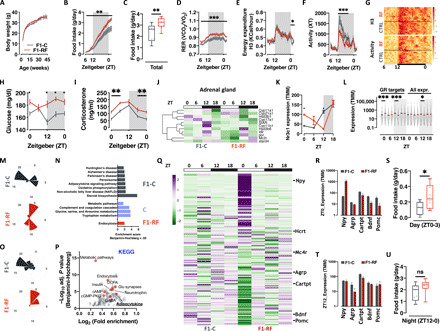
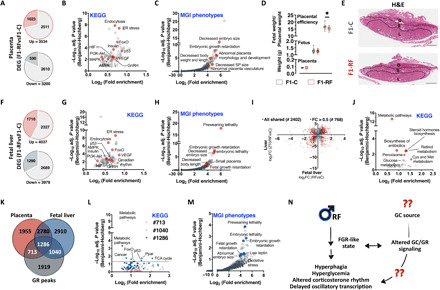
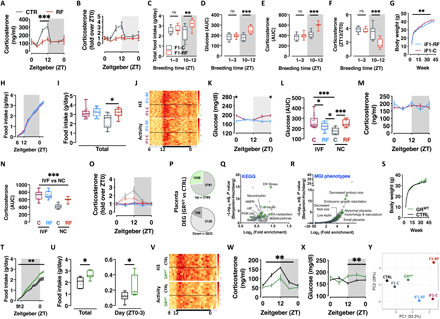
Circadian rhythm synchronizes each body function with the environment and regulates physiology. Disruption of normal circadian rhythm alters organismal physiology and increases disease risk. Recent epidemiological data and studies in model organisms have shown that maternal circadian disruption is important for offspring health and adult phenotypes. Less is known about the role of paternal circadian rhythm for offspring health. Here, we disrupted circadian rhythm in male mice by night-restricted feeding and showed that paternal circadian disruption at conception is important for offspring feeding behavior, metabolic health, and oscillatory transcription. Mechanistically, our data suggest that the effect of paternal circadian disruption is not transferred to the offspring via the germ cells but initiated by corticosterone-based parental communication at conception and programmed during in utero development through a state of fetal growth restriction. These findings indicate paternal circadian health at conception as a newly identified determinant of offspring phenotypes.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution License 4.0 (CC BY).
Figures
Comment in
-
Paternal circadian disruption affects offspring health.Lab Anim (NY). 2021 Jul;50(7):165. doi: 10.1038/s41684-021-00797-w. Lab Anim (NY). 2021. PMID: 34135532 No abstract available.
References
-
- Pittendrigh C. S., Temporal organization: Reflections of a Darwinian clock-watcher. Annu. Rev. Physiol. 55, 17–54 (1993). - PubMed
-
- Dibner C., Schibler U., Albrecht U., The mammalian circadian timing system: Organization and coordination of central and peripheral clocks. Annu. Rev. Physiol. 72, 517–549 (2010). - PubMed
-
- Golombek D. A., Rosenstein R. E., Physiology of circadian entrainment. Physiol. Rev. 90, 1063–1102 (2010). - PubMed
-
- Yoo S.-H., Yamazaki S., Lowrey P. L., Shimomura K., Ko C. H., Buhr E. D., Siepka S. M., Hong H.-K., Oh W. J., Yoo O. J., Menaker M., Takahashi J. S., PERIOD2::LUCIFERASE real-time reporting of circadian dynamics reveals persistent circadian oscillations in mouse peripheral tissues. Proc. Natl. Acad. Sci. U.S.A. 101, 5339–5346 (2004). - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
