

Novel human neutralizing mAbs specific for Spike-RBD of SARS-CoV-2
- PMID: 34040046
- PMCID: PMC8155001
- DOI: 10.1038/s41598-021-90348-7
Novel human neutralizing mAbs specific for Spike-RBD of SARS-CoV-2
Abstract
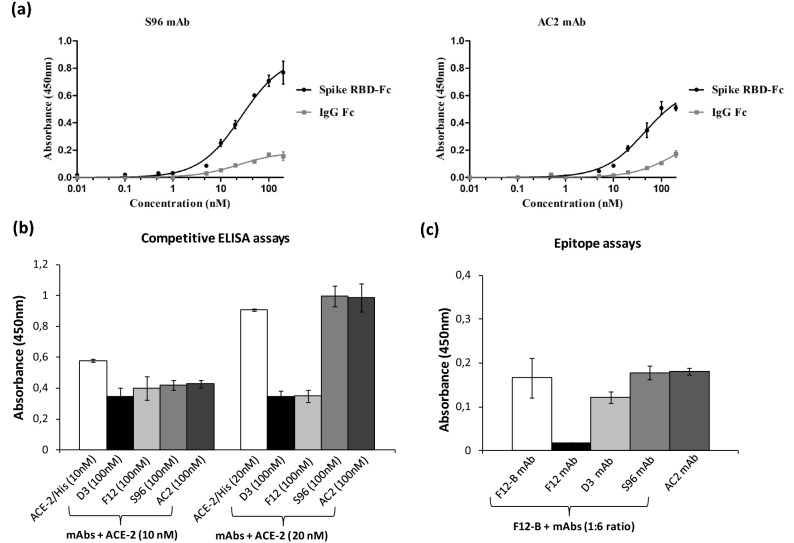
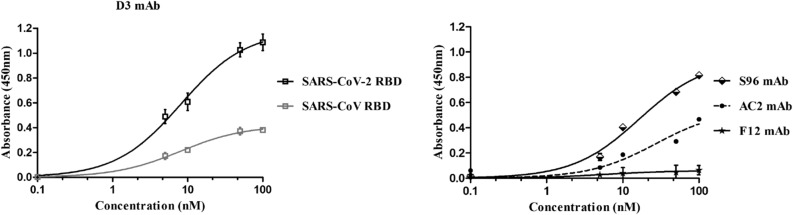
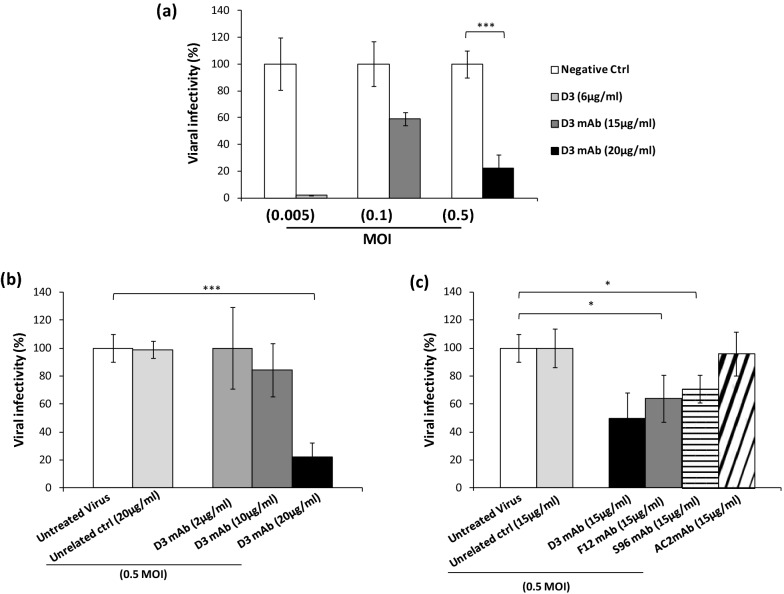
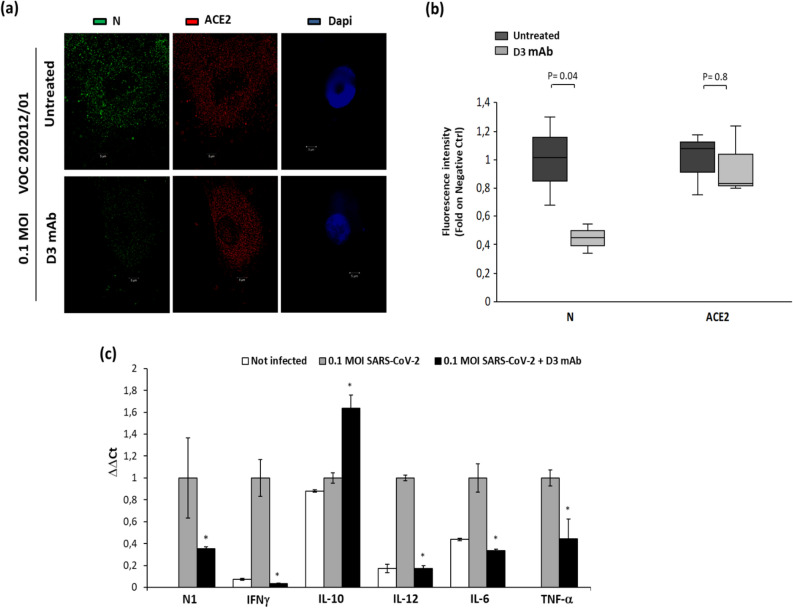
Among the therapies against the pandemic SARS-CoV-2 virus, monoclonal Antibodies (mAbs) targeting the Spike glycoprotein represent good candidates to interfere in the Spike/ACE2 interaction, preventing virus cell entry. Since anti-spike mAbs, used individually, might be unable to block the virus entry in the case of resistant mutations, we designed an innovative strategy for the isolation of multiple novel human scFvs specific for the binding domain (RBD) of Spike. By panning a large phage display antibody library on immobilized RBD, we obtained specific binders by eluting with ACE2 in order to identify those scFvs recognizing the epitope of Spike interacting with its receptor. We converted the novel scFvs into full size IgG4, differently from the previously isolated IgG1 mAbs, to avoid unwanted potential side effects of IgG1 potent effector functions on immune system. The novel antibodies specifically bind to RBD in a nanomolar range and interfere in the interaction of Spike with ACE2 receptor, either used as purified protein or when expressed on cells in its native conformation. Furthermore, some of them have neutralizing activity for virus infection in cell cultures by using two different SARS-CoV-2 isolates including the highly contagious VOC 202012/01 variant and could become useful therapeutic tools to fight against the SARS-CoV-2 virus.
Conflict of interest statement
The authors declare that a patent relative to some of the novel antibodies, mentioned in the manuscript, has been recently filed.
Figures
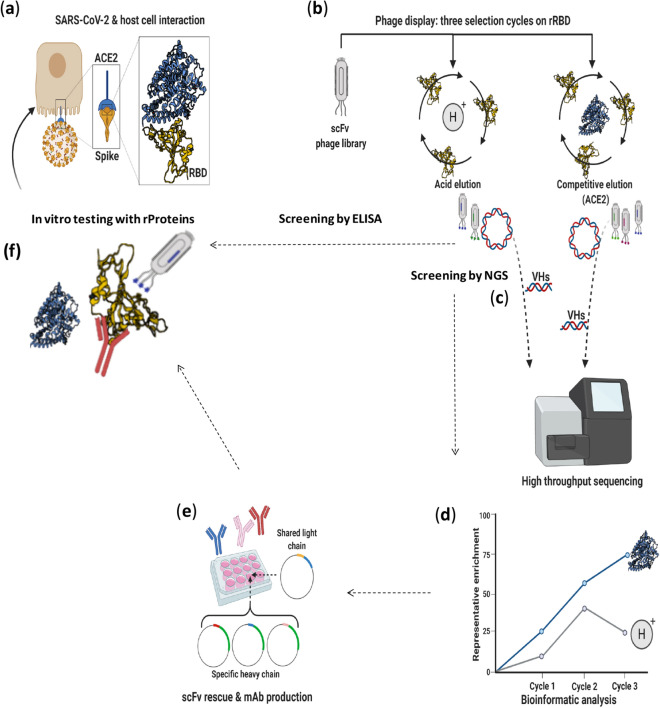
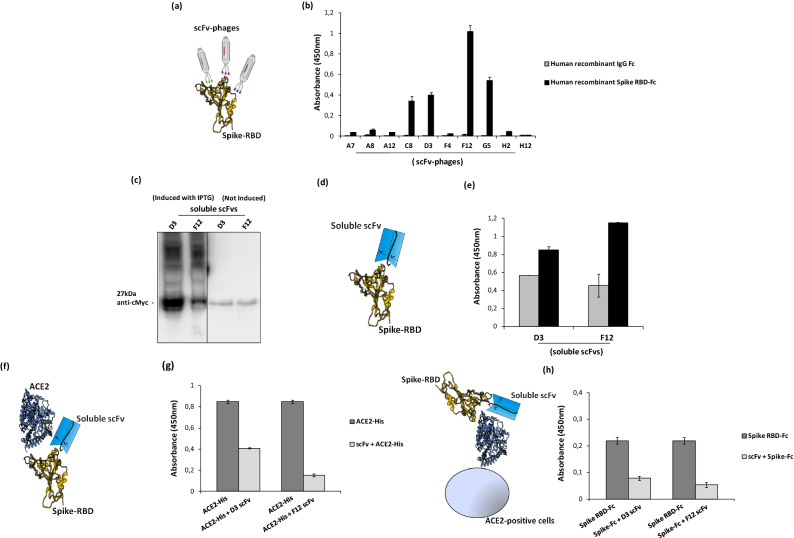
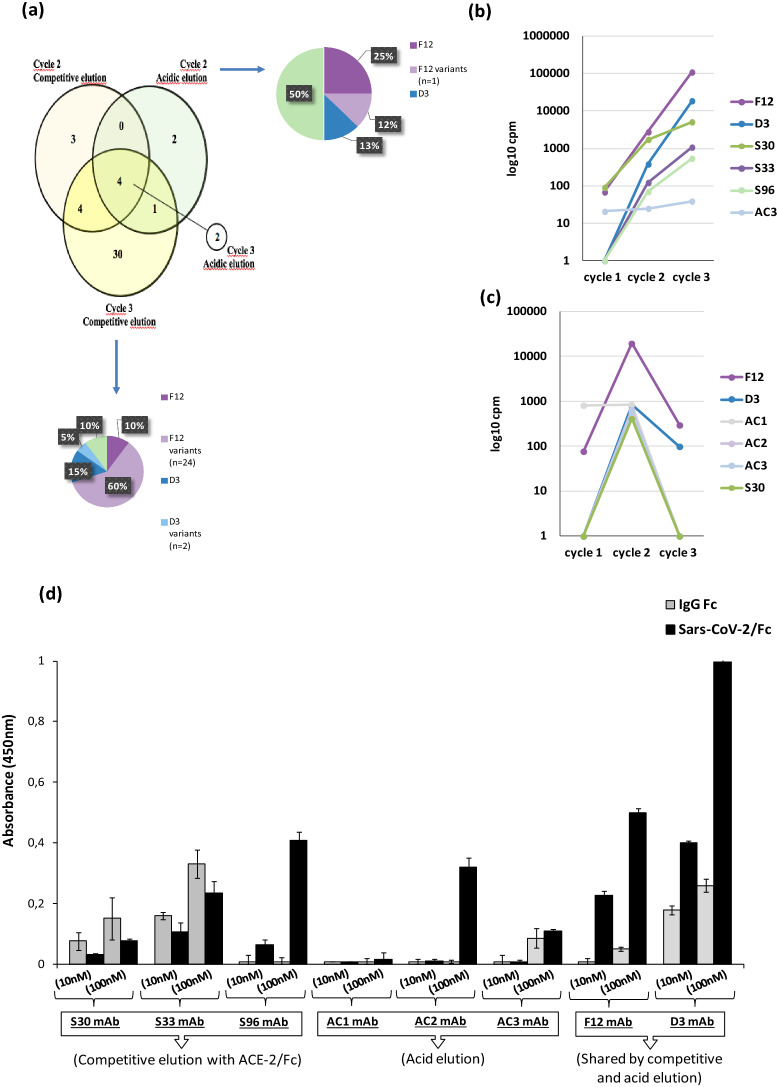
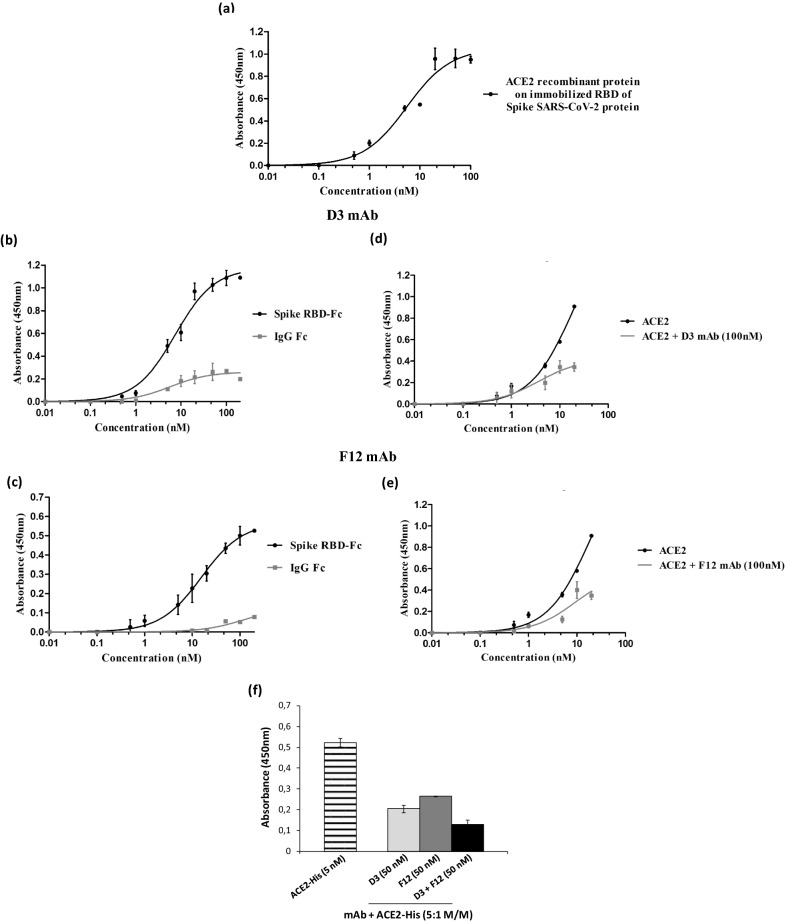
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
