

Thermally tolerant intertidal triplefin fish (Tripterygiidae) sustain ATP dynamics better than subtidal species under acute heat stress
- PMID: 34040122
- PMCID: PMC8155050
- DOI: 10.1038/s41598-021-90575-y
Thermally tolerant intertidal triplefin fish (Tripterygiidae) sustain ATP dynamics better than subtidal species under acute heat stress
Abstract
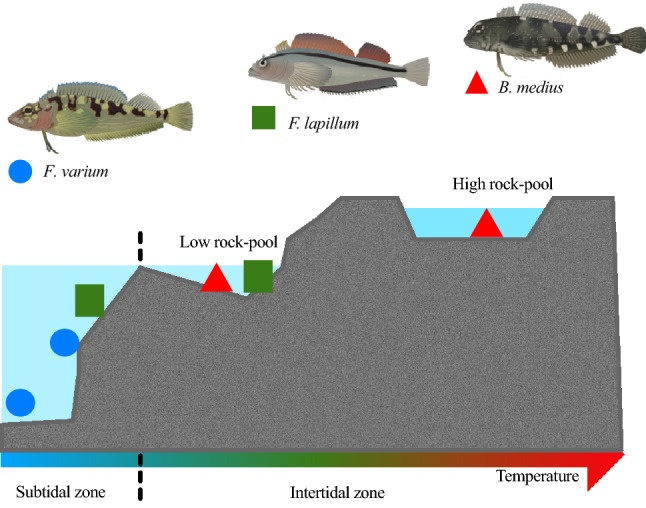
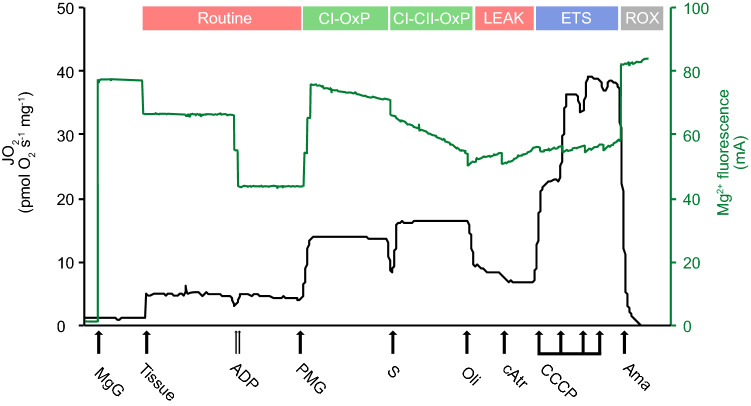
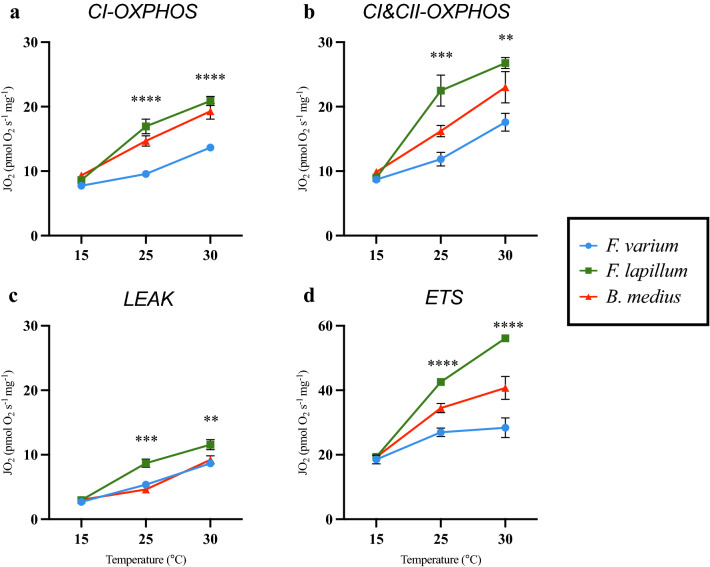
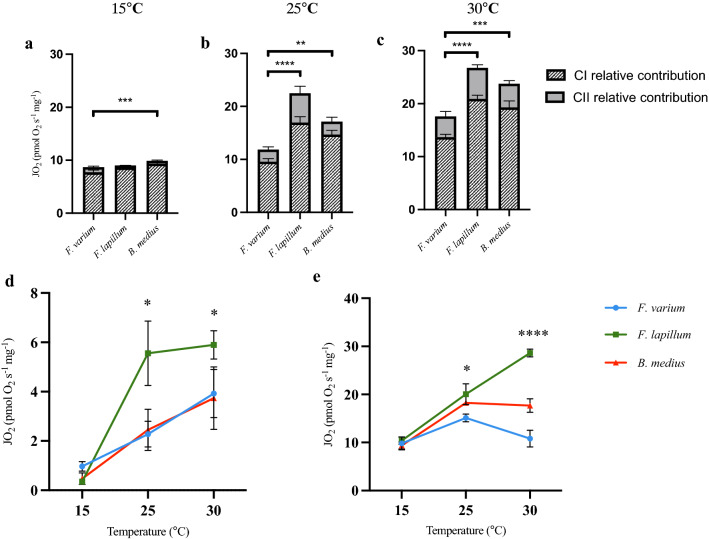
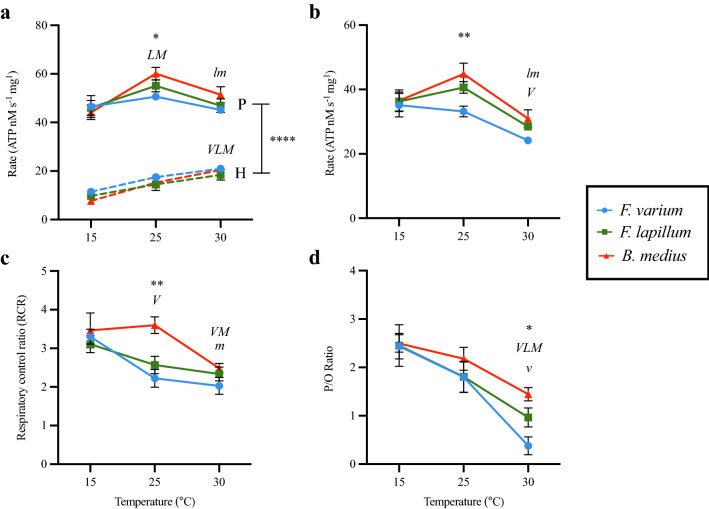
Temperature is a key factor that affects all levels of organization. Minute shifts away from thermal optima result in detrimental effects that impact growth, reproduction and survival. Metabolic rates of ectotherms are especially sensitive to temperature and for organisms exposed to high acute temperature changes, in particular intertidal species, energetic processes are often negatively impacted. Previous investigations exploring acute heat stress have implicated cardiac mitochondrial function in determining thermal tolerance. The brain, however, is by weight, one of the most metabolically active and arguably the most temperature sensitive organ. It is essentially aerobic and entirely reliant on oxidative phosphorylation to meet energetic demands, and as temperatures rise, mitochondria become less efficient at synthesising the amount of ATP required to meet the increasing demands. This leads to an energetic crisis. Here we used brain homogenate of three closely related triplefin fish species (Bellapiscis medius, Forsterygion lapillum, and Forsterygion varium) and measured respiration and ATP dynamics at three temperatures (15, 25 and 30 °C). We found that the intertidal B. medius and F. lapillum were able to maintain rates of ATP production above rates of ATP hydrolysis at high temperatures, compared to the subtidal F. varium, which showed no difference in rates at 30 °C. These results showed that brain mitochondria became less efficient at temperatures below their respective species thermal limits, and that energetic surplus of ATP synthesis over hydrolysis narrows. In subtidal species synthesis matches hydrolysis, leaving no scope to elevate ATP supply.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Thermal plasticity of skeletal muscle mitochondrial activity and whole animal respiration in a common intertidal triplefin fish, Forsterygion lapillum (Family: Tripterygiidae).J Comp Physiol B. 2014 Dec;184(8):991-1001. doi: 10.1007/s00360-014-0861-9. Epub 2014 Oct 1. J Comp Physiol B. 2014. PMID: 25269418
-
Dynamic defence? Intertidal triplefin species show better maintenance of mitochondrial membrane potential than subtidal species at low oxygen pressures.J Exp Biol. 2023 Aug 15;226(16):jeb245926. doi: 10.1242/jeb.245926. Epub 2023 Aug 22. J Exp Biol. 2023. PMID: 37498237
-
Temperature sensitivity of cardiac mitochondria in intertidal and subtidal triplefin fishes.J Comp Physiol B. 2010 Oct;180(7):979-90. doi: 10.1007/s00360-010-0477-7. Epub 2010 May 12. J Comp Physiol B. 2010. PMID: 20461387
-
Variation in the heat shock response and its implication for predicting the effect of global climate change on species' biogeographical distribution ranges and metabolic costs.J Exp Biol. 2010 Mar 15;213(6):971-9. doi: 10.1242/jeb.038034. J Exp Biol. 2010. PMID: 20190122 Review.
-
Cellular energy utilization and molecular origin of standard metabolic rate in mammals.Physiol Rev. 1997 Jul;77(3):731-58. doi: 10.1152/physrev.1997.77.3.731. Physiol Rev. 1997. PMID: 9234964 Review.
Cited by
-
Ecophysiological responses to heat waves in the marine intertidal zone.J Exp Biol. 2025 Jan 15;228(2):JEB246503. doi: 10.1242/jeb.246503. Epub 2025 Jan 16. J Exp Biol. 2025. PMID: 39817480 Free PMC article. Review.
References
-
- Hochachka PW, Somero GN. Biochemical Adaptation: Mechanism and Process in Physiological Evolution. Oxford University Press; 2002.
-
- Helmuth B, et al. Living on the Edge of Two Changing Worlds: Forecasting the Responses of Rocky Intertidal Ecosystems to Climate Change. ECU Publications; 2006.
-
- Woodward A. Climate change: Disruption, risk and opportunity. Glob. Transit. 2019;1:44–49. doi: 10.1016/j.glt.2019.02.001. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
