

NAD+ homeostasis in human health and disease
- PMID: 34041853
- PMCID: PMC8261484
- DOI: 10.15252/emmm.202113943
NAD+ homeostasis in human health and disease
Abstract
Depletion of nicotinamide adenine dinucleotide (NAD+ ), a central redox cofactor and the substrate of key metabolic enzymes, is the causative factor of a number of inherited and acquired diseases in humans. Primary deficiencies of NAD+ homeostasis are the result of impaired biosynthesis, while secondary deficiencies can arise due to other factors affecting NAD+ homeostasis, such as increased NAD+ consumption or dietary deficiency of its vitamin B3 precursors. NAD+ depletion can manifest in a wide variety of pathological phenotypes, ranging from rare inherited defects, characterized by congenital malformations, retinal degeneration, and/or encephalopathy, to more common multifactorial, often age-related, diseases. Here, we discuss NAD+ biochemistry and metabolism and provide an overview of the etiology and pathological consequences of alterations of the NAD+ metabolism in humans. Finally, we discuss the state of the art of the potential therapeutic implications of NAD+ repletion for boosting health as well as treating rare and common diseases, and the possibilities to achieve this by means of the different NAD+ -enhancing agents.
Keywords: NAD+; NAD+ homeostasis; disease; metabolism; therapy.
© 2021 The Authors. Published under the terms of the CC BY 4.0 license.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
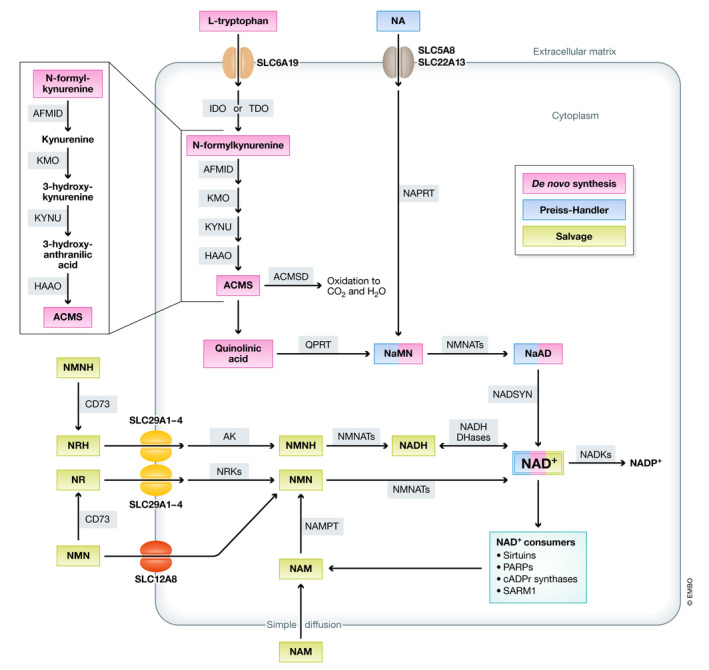
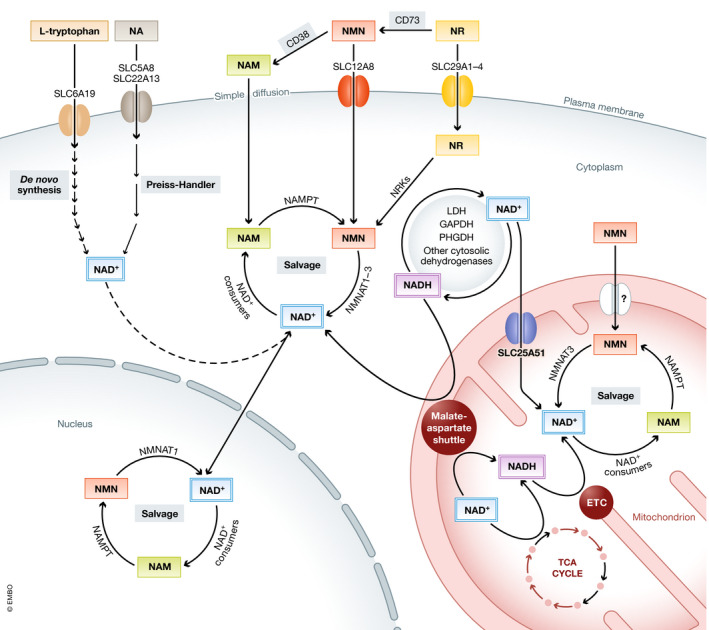
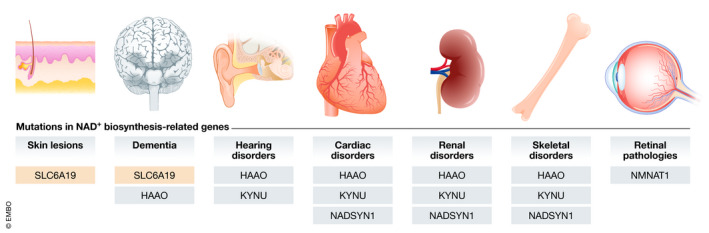
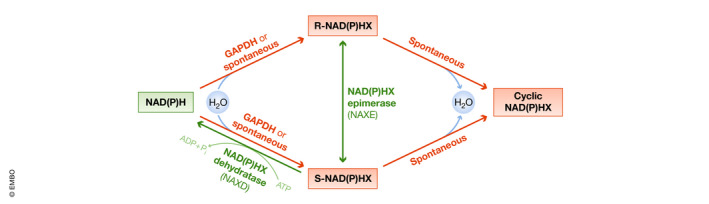
References
-
- Airhart SE, Shireman LM, Risler LJ, Anderson GD, Nagana Gowda GA, Raftery D, Tian R, Shen DD, O'Brien KD (2017) An open‐label, non‐randomized study of the pharmacokinetics of the nutritional supplement nicotinamide riboside (NR) and its effects on blood NAD+ levels in healthy volunteers. PLoS One 12: e0186459 - PMC - PubMed
-
- Aksoy P, White TA, Thompson M, Chini EN (2006) Regulation of intracellular levels of NAD: a novel role for CD38. Biochem Biophys Res Commun 345: 1386–1392 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
