

Regulation of Perineuronal Nets in the Adult Cortex by the Activity of the Cortical Network
- PMID: 34045309
- PMCID: PMC8265812
- DOI: 10.1523/JNEUROSCI.0434-21.2021
Regulation of Perineuronal Nets in the Adult Cortex by the Activity of the Cortical Network
Abstract
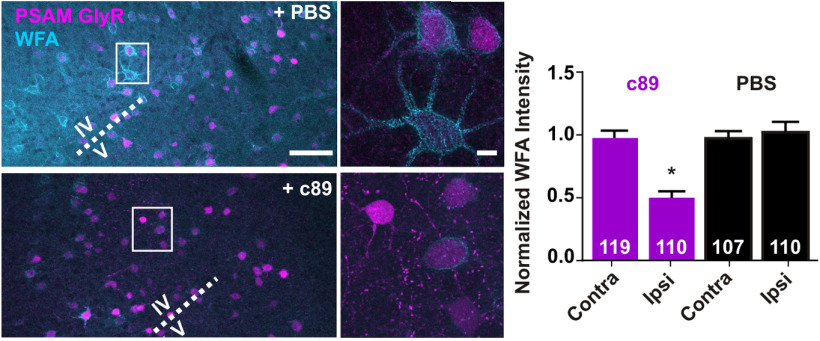
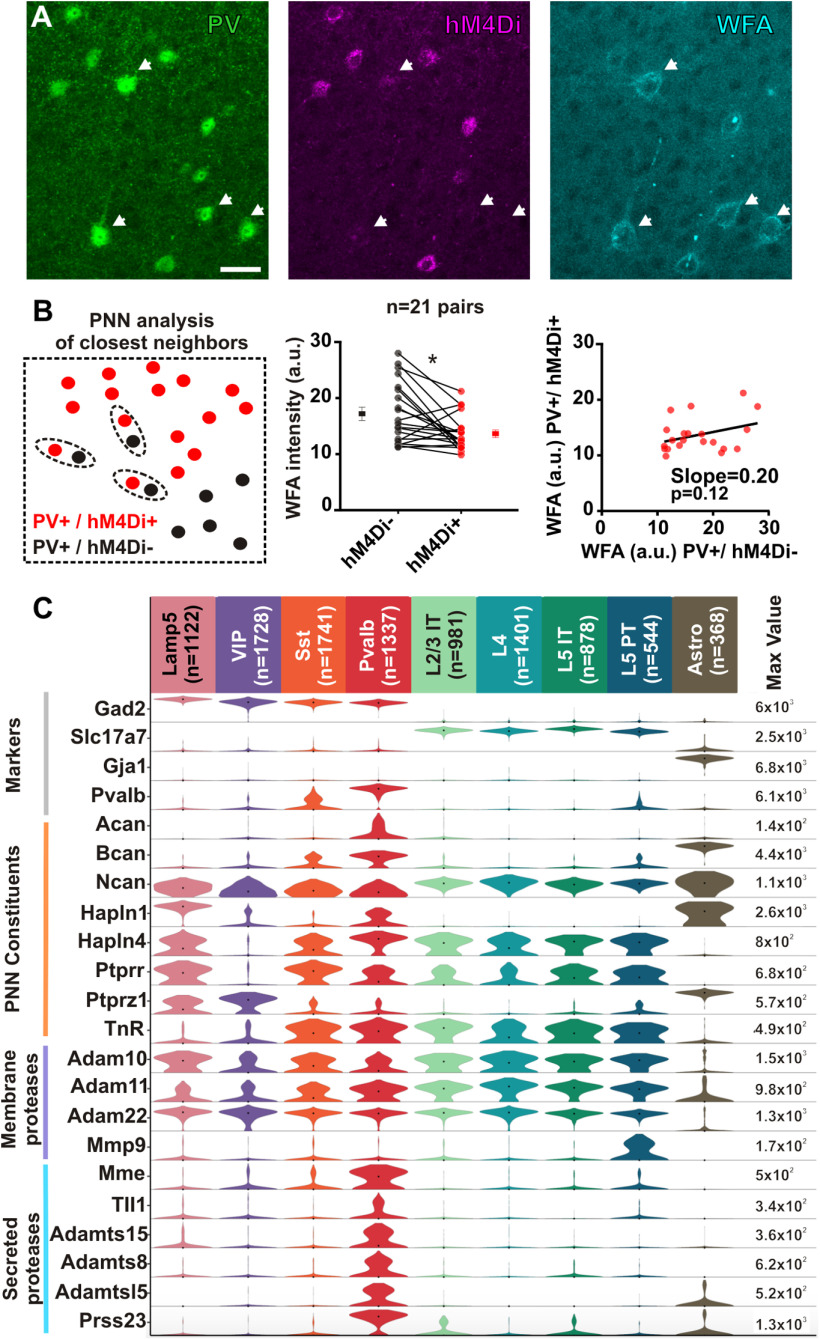
Perineuronal net (PNN) accumulation around parvalbumin-expressing (PV) inhibitory interneurons marks the closure of critical periods of high plasticity, whereas PNN removal reinstates juvenile plasticity in the adult cortex. Using targeted chemogenetic in vivo approaches in the adult mouse visual cortex, we found that transient inhibition of PV interneurons, through metabotropic or ionotropic chemogenetic tools, induced PNN regression. EEG recordings indicated that inhibition of PV interneurons did not elicit unbalanced network excitation. Likewise, inhibition of local excitatory neurons also induced PNN regression, whereas chemogenetic excitation of either PV or excitatory neurons did not reduce the PNN. We also observed that chemogenetically inhibited PV interneurons exhibited reduced PNN compared with their untransduced neighbors, and confirmed that single PV interneurons express multiple genes enabling individual regulation of their own PNN density. Our results indicate that PNN density is regulated in the adult cortex by local changes of network activity that can be triggered by modulation of PV interneurons. PNN regulation may provide adult cortical circuits with an activity-dependent mechanism to control their local remodeling.SIGNIFICANCE STATEMENT The perineuronal net is an extracellular matrix, which accumulates around individual parvalbumin-expressing inhibitory neurons during postnatal development, and is seen as a barrier that prevents plasticity of neuronal circuits in the adult cerebral cortex. We found that transiently inhibiting parvalbumin-expressing or excitatory cortical neurons triggers a local decrease of perineuronal net density. Our results indicate that perineuronal nets are regulated in the adult cortex depending on the activity of local microcircuits. These findings uncover an activity-dependent mechanism by which adult cortical circuits may locally control their plasticity.
Keywords: cerebral cortex; critical period plasticity; extracellular matrix; fast-spiking parvalbumin interneurons; perineuronal net.
Copyright © 2021 the authors.
Figures






Similar articles
-
Experience-dependent development of perineuronal nets and chondroitin sulfate proteoglycan receptors in mouse visual cortex.Matrix Biol. 2013 Aug 8;32(6):352-63. doi: 10.1016/j.matbio.2013.04.001. Epub 2013 Apr 15. Matrix Biol. 2013. PMID: 23597636
-
Effects of noise-induced hearing loss on parvalbumin and perineuronal net expression in the mouse primary auditory cortex.Hear Res. 2017 Jul;350:82-90. doi: 10.1016/j.heares.2017.04.015. Epub 2017 Apr 27. Hear Res. 2017. PMID: 28460252
-
Removal of Perineuronal Nets Unlocks Juvenile Plasticity Through Network Mechanisms of Decreased Inhibition and Increased Gamma Activity.J Neurosci. 2017 Feb 1;37(5):1269-1283. doi: 10.1523/JNEUROSCI.2504-16.2016. Epub 2016 Dec 30. J Neurosci. 2017. PMID: 28039374 Free PMC article.
-
The Perineuronal 'Safety' Net? Perineuronal Net Abnormalities in Neurological Disorders.Front Mol Neurosci. 2018 Aug 3;11:270. doi: 10.3389/fnmol.2018.00270. eCollection 2018. Front Mol Neurosci. 2018. PMID: 30123106 Free PMC article. Review.
-
A type-5 metabotropic glutamate receptor-perineuronal net axis shapes the function of cortical GABAergic interneurons in chronic pain.J Anesth Analg Crit Care. 2025 Feb 21;5(1):10. doi: 10.1186/s44158-025-00228-z. J Anesth Analg Crit Care. 2025. PMID: 39985105 Free PMC article. Review.
Cited by
-
Parvalbumin - Positive Neurons in the Neocortex: A Review.Physiol Res. 2023 Jul 31;72(Suppl 2):S173-S191. doi: 10.33549/physiolres.935005. Physiol Res. 2023. PMID: 37565421 Free PMC article. Review.
-
Metabotropic signaling within somatostatin interneurons controls transient thalamocortical inputs during development.Nat Commun. 2024 Jun 26;15(1):5421. doi: 10.1038/s41467-024-49732-w. Nat Commun. 2024. PMID: 38926335 Free PMC article.
-
EHMT1 regulates Parvalbumin-positive interneuron development and GABAergic input in sensory cortical areas.Brain Struct Funct. 2020 Dec;225(9):2701-2716. doi: 10.1007/s00429-020-02149-9. Epub 2020 Sep 25. Brain Struct Funct. 2020. PMID: 32975655 Free PMC article.
-
The Perineuronal Net Protein Brevican Acts in Nucleus Accumbens Parvalbumin-Expressing Interneurons of Adult Mice to Regulate Excitatory Synaptic Inputs and Motivated Behaviors.Biol Psychiatry. 2024 Nov 1;96(9):694-707. doi: 10.1016/j.biopsych.2024.02.003. Epub 2024 Feb 10. Biol Psychiatry. 2024. PMID: 38346480
-
A sensitive period for the development of episodic-like memory in mice.bioRxiv [Preprint]. 2024 Nov 7:2024.11.06.622296. doi: 10.1101/2024.11.06.622296. bioRxiv. 2024. Update in: Curr Biol. 2025 May 5;35(9):2032-2048.e3. doi: 10.1016/j.cub.2025.03.032. PMID: 39574753 Free PMC article. Updated. Preprint.
References
-
- Alexander GM, Rogan SC, Abbas AI, Armbruster BN, Pei Y, Allen JA, Nonneman RJ, Hartmann J, Moy SS, Nicolelis MA, McNamara JO, Roth BL (2009) Remote control of neuronal activity in transgenic mice expressing evolved G protein-coupled receptors. Neuron 63:27–39. 10.1016/j.neuron.2009.06.014 - DOI - PMC - PubMed
-
- Ascoli GA, Alonso-Nanclares L, Anderson SA, Barrionuevo G, Benavides-Piccione R, Burkhalter A, Buzsáki G, Cauli B, Defelipe J, Fairén A, Feldmeyer D, Fishell G, Fregnac Y, Freund TF, Gardner D, Gardner EP, Goldberg JH, Helmstaedter M, Hestrin S, Karube F, et al. (2008) Petilla terminology: nomenclature of features of GABAergic interneurons of the cerebral cortex. Nat Rev Neurosci 9:557–568. 10.1038/nrn2402 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials