

Single tryptophan Y160W mutant of homooligomeric E. coli purine nucleoside phosphorylase implies that dimers forming the hexamer are functionally not equivalent
- PMID: 34045551
- PMCID: PMC8160210
- DOI: 10.1038/s41598-021-90472-4
Single tryptophan Y160W mutant of homooligomeric E. coli purine nucleoside phosphorylase implies that dimers forming the hexamer are functionally not equivalent
Abstract
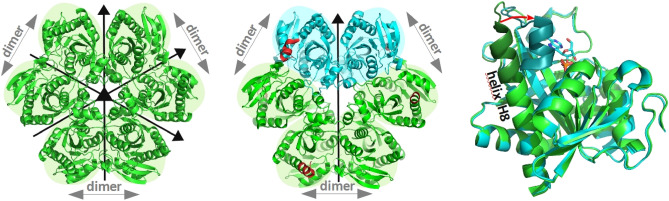
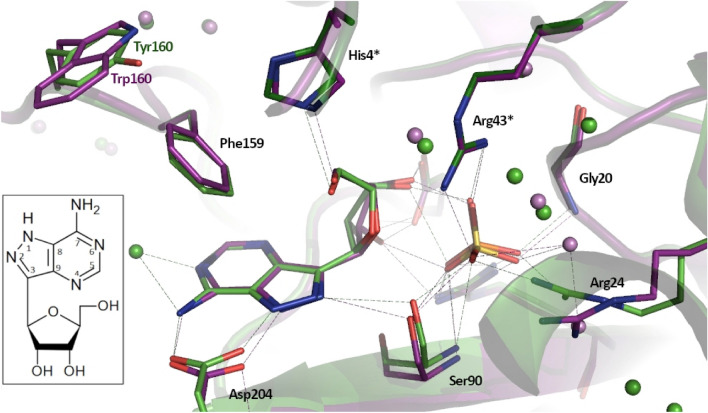
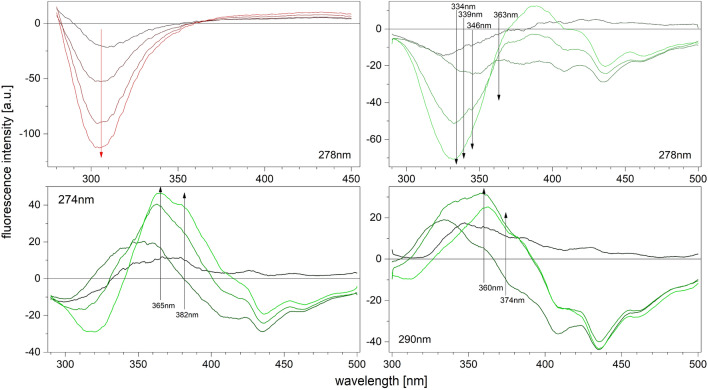
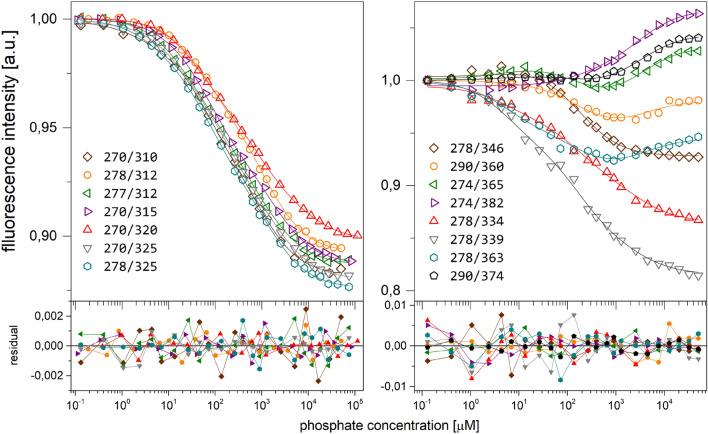
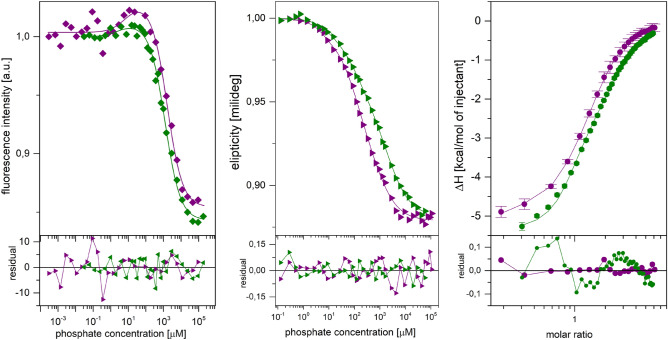
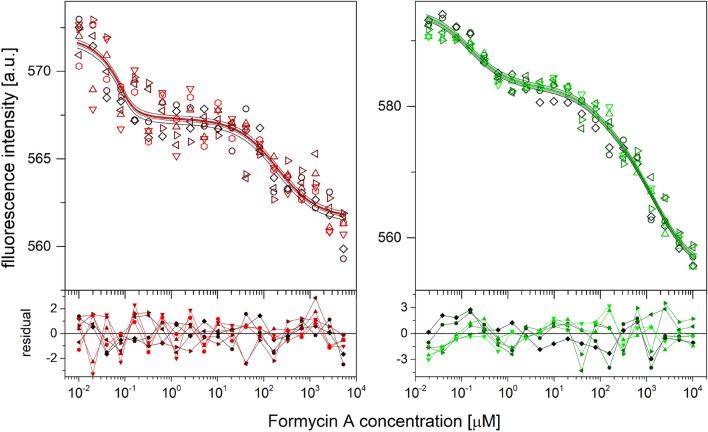
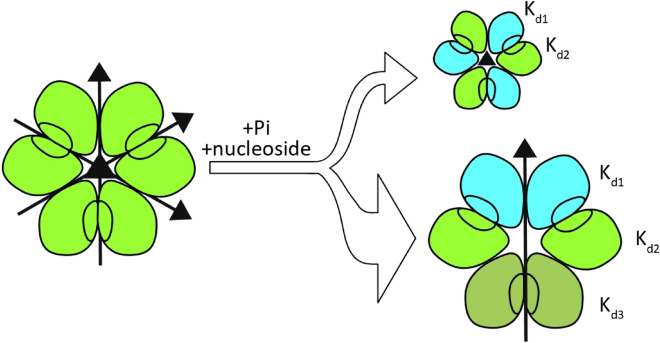
E. coli purine nucleoside phosphorylase is a homohexamer, which structure, in the apo form, can be described as a trimer of dimers. Earlier studies suggested that ligand binding and kinetic properties are well described by two binding constants and two sets of kinetic constants. However, most of the crystal structures of this enzyme complexes with ligands do not hold the three-fold symmetry, but only two-fold symmetry, as one of the three dimers is different (both active sites in the open conformation) from the other two (one active site in the open and one in the closed conformation). Our recent detailed studies conducted over broad ligand concentration range suggest that protein-ligand complex formation in solution actually deviates from the two-binding-site model. To reveal the details of interactions present in the hexameric molecule we have engineered a single tryptophan Y160W mutant, responding with substantial intrinsic fluorescence change upon ligand binding. By observing various physical properties of the protein and its various complexes with substrate and substrate analogues we have shown that indeed three-binding-site model is necessary to properly describe binding of ligands by both the wild type enzyme and the Y160W mutant. Thus we have pointed out that a symmetrical dimer with both active sites in the open conformation is not forced to adopt this conformation by interactions in the crystal, but most probably the dimers forming the hexamer in solution are not equivalent as well. This, in turn, implies that an allosteric cooperation occurs not only within a dimer, but also among all three dimers forming a hexameric molecule.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Crystallographic snapshots of ligand binding to hexameric purine nucleoside phosphorylase and kinetic studies give insight into the mechanism of catalysis.Sci Rep. 2018 Oct 18;8(1):15427. doi: 10.1038/s41598-018-33723-1. Sci Rep. 2018. PMID: 30337572 Free PMC article.
-
Open and closed conformation of the E. coli purine nucleoside phosphorylase active center and implications for the catalytic mechanism.J Mol Biol. 2002 Jan 18;315(3):351-71. doi: 10.1006/jmbi.2001.5211. J Mol Biol. 2002. PMID: 11786017
-
Homooligomerization is needed for stability: a molecular modelling and solution study of Escherichia coli purine nucleoside phosphorylase.FEBS J. 2014 Apr;281(7):1860-71. doi: 10.1111/febs.12746. FEBS J. 2014. PMID: 24785777
-
New Insights into Active Site Conformation Dynamics of E. coli PNP Revealed by Combined H/D Exchange Approach and Molecular Dynamics Simulations.J Am Soc Mass Spectrom. 2016 Jan;27(1):73-82. doi: 10.1007/s13361-015-1239-2. Epub 2015 Sep 3. J Am Soc Mass Spectrom. 2016. PMID: 26337516
-
Structural analyses reveal two distinct families of nucleoside phosphorylases.Biochem J. 2002 Jan 1;361(Pt 1):1-25. doi: 10.1042/0264-6021:3610001. Biochem J. 2002. PMID: 11743878 Free PMC article. Review.
Cited by
-
Thermodynamic Studies of Interactions between Sertraline Hydrochloride and Randomly Methylated β-Cyclodextrin Molecules Supported by Circular Dichroism Spectroscopy and Molecular Docking Results.Int J Mol Sci. 2021 Nov 16;22(22):12357. doi: 10.3390/ijms222212357. Int J Mol Sci. 2021. PMID: 34830239 Free PMC article.
-
Differential scanning fluorimetry followed by microscale thermophoresis and/or isothermal titration calorimetry as an efficient tool for ligand screening.Biophys Rev. 2025 Feb 13;17(1):199-223. doi: 10.1007/s12551-025-01280-3. eCollection 2025 Feb. Biophys Rev. 2025. PMID: 40060009 Free PMC article. Review.
-
The structure of His-tagged Geobacillus stearothermophilus purine nucleoside phosphorylase reveals a `spanner in the works'.Acta Crystallogr F Struct Biol Commun. 2022 Dec 1;78(Pt 12):416-422. doi: 10.1107/S2053230X22011025. Epub 2022 Nov 28. Acta Crystallogr F Struct Biol Commun. 2022. PMID: 36458621 Free PMC article.
-
Interaction of Tri-Cyclic Nucleobase Analogs with Enzymes of Purine Metabolism: Xanthine Oxidase and Purine Nucleoside Phosphorylase.Int J Mol Sci. 2024 Sep 27;25(19):10426. doi: 10.3390/ijms251910426. Int J Mol Sci. 2024. PMID: 39408755 Free PMC article.
-
The Interaction of Heptakis (2,6-di-O-Methyl)-β-cyclodextrin with Mianserin Hydrochloride and Its Influence on the Drug Toxicity.Int J Mol Sci. 2021 Aug 30;22(17):9419. doi: 10.3390/ijms22179419. Int J Mol Sci. 2021. PMID: 34502332 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
