

Contrasting roles of GmNAC065 and GmNAC085 in natural senescence, plant development, multiple stresses and cell death responses
- PMID: 34045652
- PMCID: PMC8160357
- DOI: 10.1038/s41598-021-90767-6
Contrasting roles of GmNAC065 and GmNAC085 in natural senescence, plant development, multiple stresses and cell death responses
Abstract
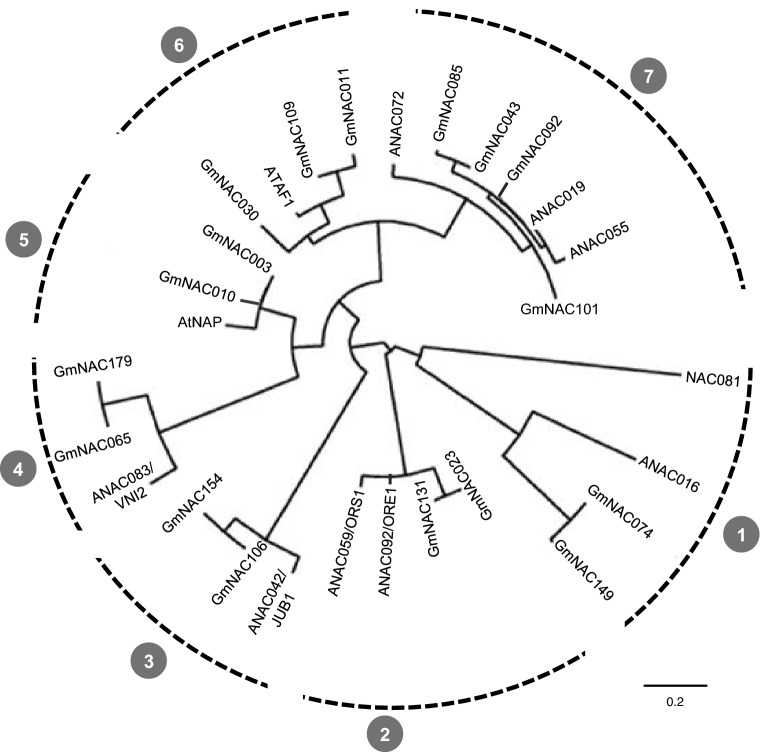
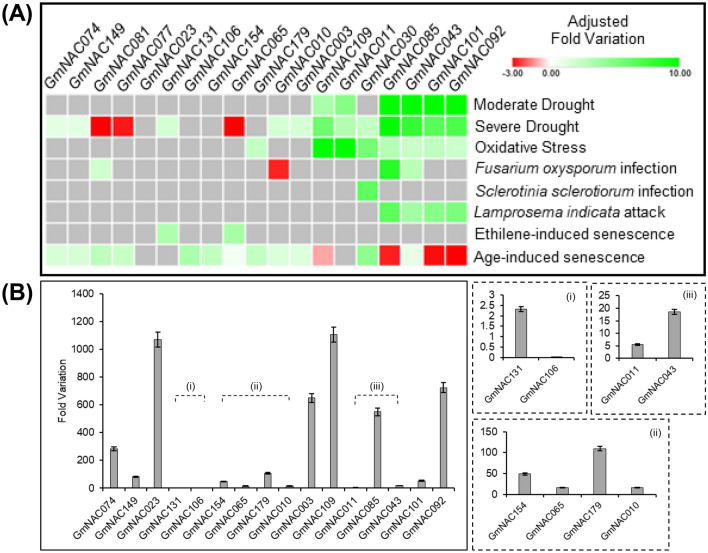
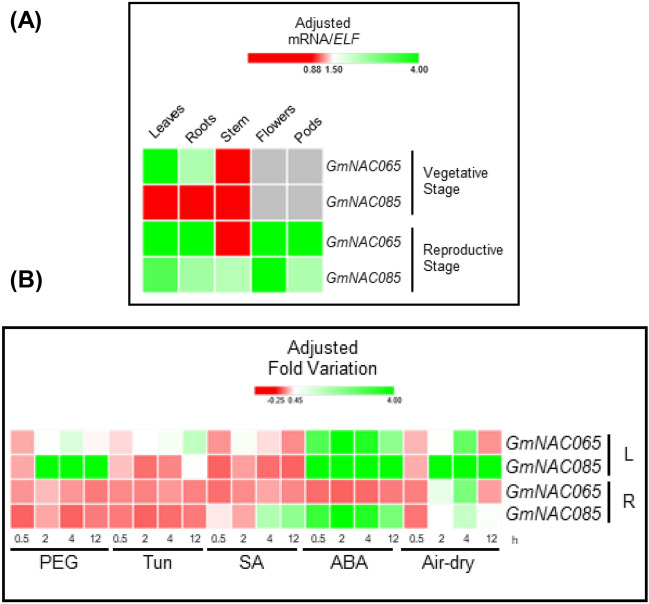
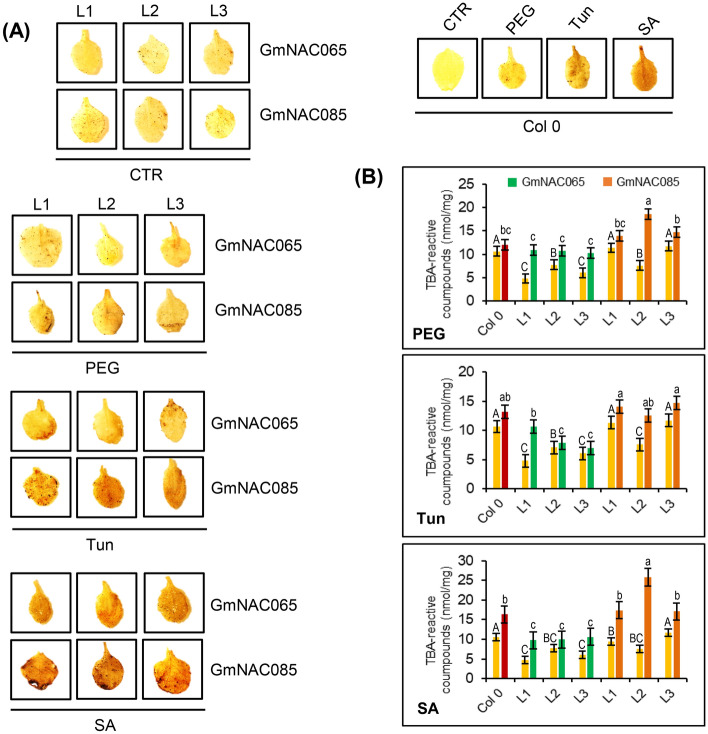
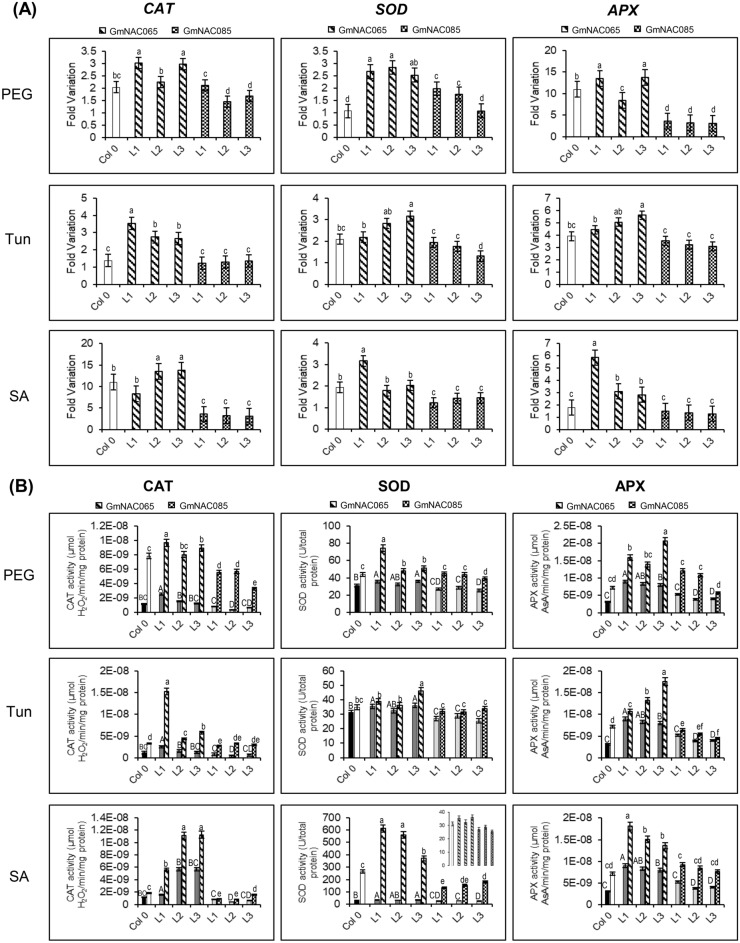
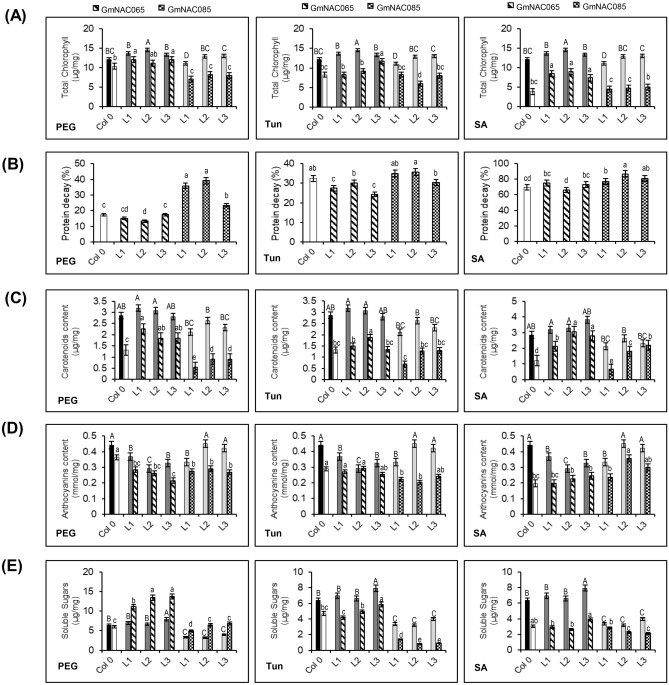
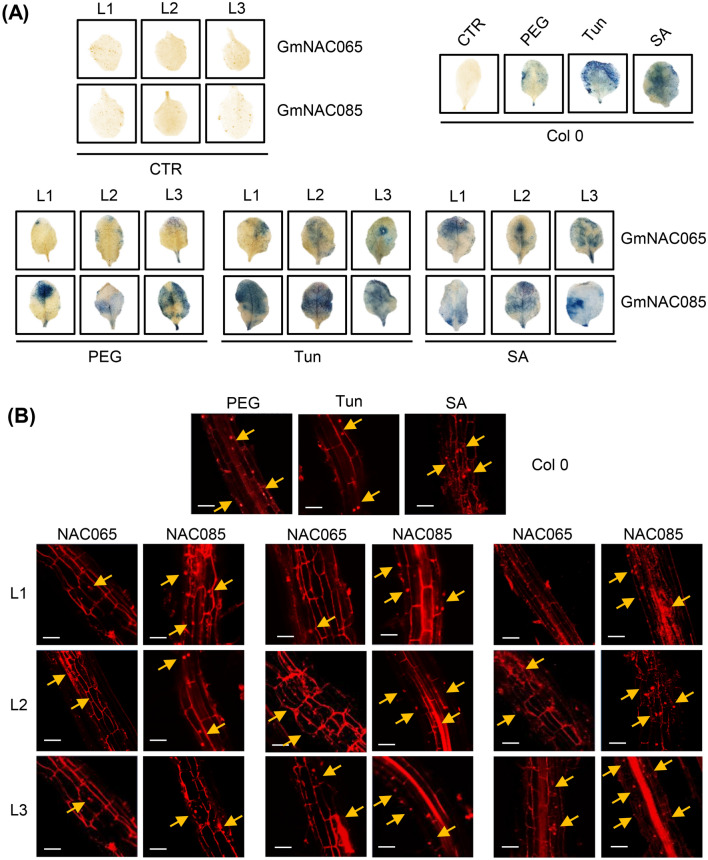
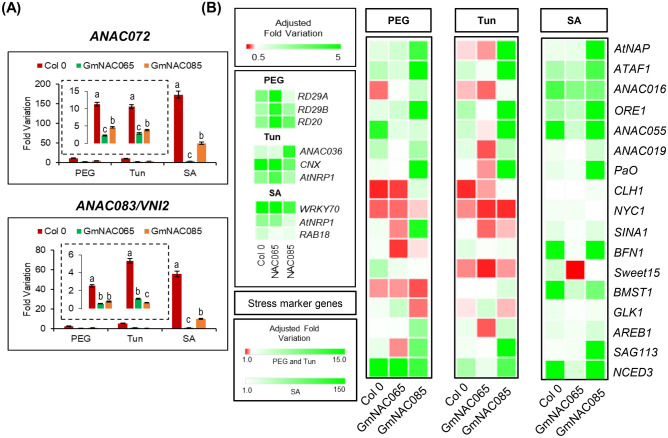
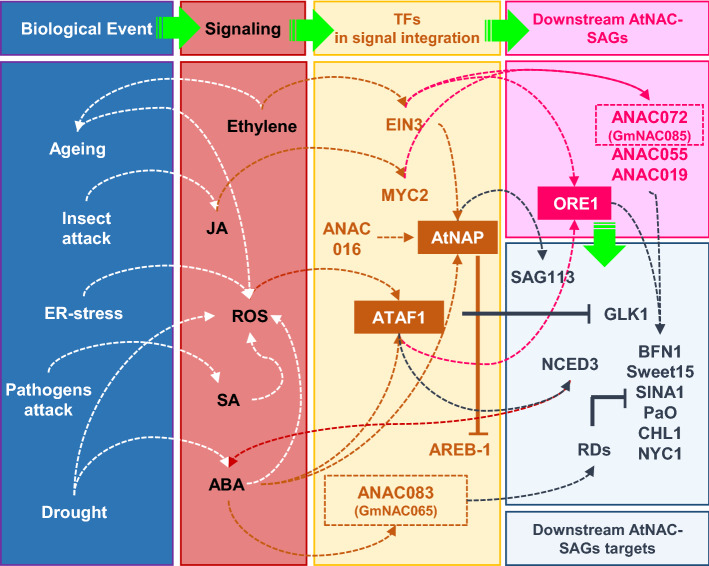
NACs are plant-specific transcription factors involved in controlling plant development, stress responses, and senescence. As senescence-associated genes (SAGs), NACs integrate age- and stress-dependent pathways that converge to programmed cell death (PCD). In Arabidopsis, NAC-SAGs belong to well-characterized regulatory networks, poorly understood in soybean. Here, we interrogated the soybean genome and provided a comprehensive analysis of senescence-associated Glycine max (Gm) NACs. To functionally examine GmNAC-SAGs, we selected GmNAC065, a putative ortholog of Arabidopsis ANAC083/VNI2 SAG, and the cell death-promoting GmNAC085, an ANAC072 SAG putative ortholog, for analyses. Expression analysis of GmNAC065 and GmNAC085 in soybean demonstrated (i) these cell death-promoting GmNACs display contrasting expression changes during age- and stress-induced senescence; (ii) they are co-expressed with functionally different gene sets involved in stress and PCD, and (iii) are differentially induced by PCD inducers. Furthermore, we demonstrated GmNAC065 expression delays senescence in Arabidopsis, a phenotype associated with enhanced oxidative performance under multiple stresses, higher chlorophyll, carotenoid and sugar contents, and lower stress-induced PCD compared to wild-type. In contrast, GmNAC085 accelerated stress-induced senescence, causing enhanced chlorophyll loss, ROS accumulation and cell death, decreased antioxidative system expression and activity. Accordingly, GmNAC065 and GmNAC085 targeted functionally contrasting sets of downstream AtSAGs, further indicating that GmNAC85 and GmNAC065 regulators function inversely in developmental and environmental PCD.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
