

Sensory adaptation at ribbon synapses in the zebrafish lateral line
- PMID: 34047358
- PMCID: PMC7612133
- DOI: 10.1113/JP281646
Sensory adaptation at ribbon synapses in the zebrafish lateral line
Abstract
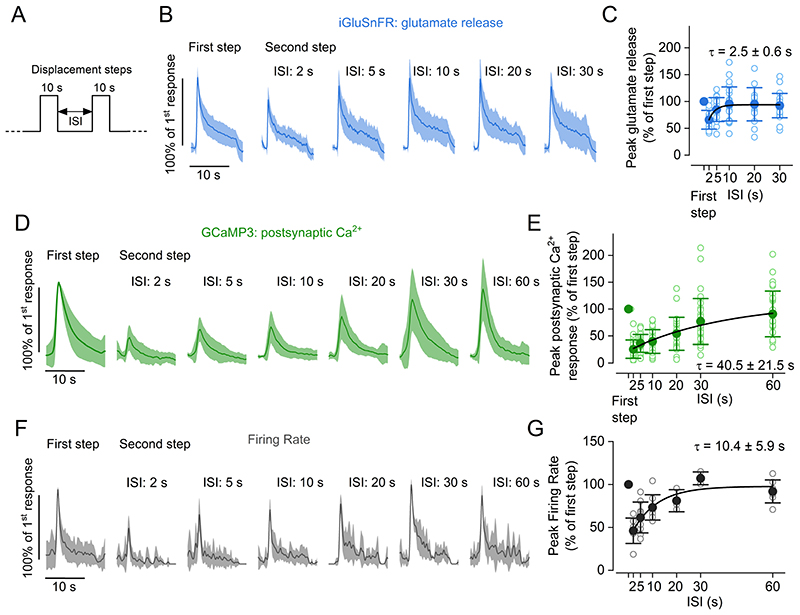
Key points: The present study aimed to determine the sensory adaptation characteristics of hair cell ribbon synapses in vivo. Hair cells of the zebrafish lateral line transmit hydrodynamic stimuli to the posterior lateral line ganglion afferent neurons. Excitatory hair bundle deflections by water-jet stimuli cause glutamate release at hair cell synapses with a rapid (phasic) and a sustained component, which are likely linked to the exocytosis of distinct vesicle pools. The glutamate-induced increase in afferent neuron firing rate adapts over time, which is mirrored by the depression of neurotransmitter release, without preventing phase-locking. Adaptation also occurs during inhibitory hair bundle displacements, highlighting a shift in the sensitivity range of the lateral line during prolonged stimulation. Postsynaptic mechanisms exert some degree of regulation on the afferent firing adaptation. We conclude that vesicle depletion is the primary determinant of firing rate adaptation, allowing lateral line hair cell ribbon synapses to maintain sensitivity to sustained stimuli.
Abstract: Adaptation is used by sensory systems to adjust continuously their sensitivity to match changes in environmental stimuli. In the auditory and vestibular systems, the release properties of glutamate-containing vesicles at the hair cell ribbon synapses play a crucial role in sensory adaptation, thus shaping the neural response to sustained stimulation. How ribbon synapses regulate the release of glutamate and how they modulate afferent responses in vivo is still largely unknown. Here, we have used two-photon imaging and electrophysiology to investigate the synaptic transfer characteristics of the hair cells in the context of sensory adaptation in live zebrafish. Prolonged and repeated water-jet stimulation of the hair cell stereociliary bundles caused adaptation of the action potential firing rate elicited in the afferent neurons. By monitoring glutamate at ribbon synapses using time-lapse imaging, we identified two kinetically distinct release components: a rapid response that was exhausted within 50-100 ms and a slower and sustained response lasting the entire stimulation. After repeated stimulations, the recovery of the fast component followed a biphasic time course. Depression of glutamate release was largely responsible for the rapid firing rate adaptation recorded in the afferent neurons. However, postsynaptic Ca2+ responses had a slower recovery time course compared to that of glutamate release, indicating that they are likely to contribute to the afferent firing adaptation. Hair cells also exhibited a form of adaptation during inhibitory bundle stimulations. We conclude that hair cells have optimised their synaptic machinery to encode prolonged stimuli and to maintain their sensitivity to new incoming stimuli.
Keywords: adaptation; hair cells; in vivo physiology; synaptic release; vesicle pools; zebrafish lateral line.
© 2021 The Authors. The Journal of Physiology published by John Wiley & Sons Ltd on behalf of The Physiological Society.
Conflict of interest statement
The Authors declare no conflict of interest.
Figures








Similar articles
-
The Transfer Characteristics of Hair Cells Encoding Mechanical Stimuli in the Lateral Line of Zebrafish.J Neurosci. 2019 Jan 2;39(1):112-124. doi: 10.1523/JNEUROSCI.1472-18.2018. Epub 2018 Nov 9. J Neurosci. 2019. PMID: 30413644 Free PMC article.
-
Functional development and regeneration of hair cells in the zebrafish lateral line.J Physiol. 2021 Aug;599(16):3913-3936. doi: 10.1113/JP281522. Epub 2021 Jul 9. J Physiol. 2021. PMID: 34143497 Free PMC article.
-
Kif1a and intact microtubules maintain synaptic-vesicle populations at ribbon synapses in zebrafish hair cells.J Physiol. 2024 Oct 7:10.1113/JP286263. doi: 10.1113/JP286263. Online ahead of print. J Physiol. 2024. PMID: 39373584
-
Structure and function of the hair cell ribbon synapse.J Membr Biol. 2006 Feb-Mar;209(2-3):153-65. doi: 10.1007/s00232-005-0854-4. Epub 2006 May 25. J Membr Biol. 2006. PMID: 16773499 Free PMC article. Review.
-
Molecular organization of a type of peripheral glutamate synapse: the afferent synapses of hair cells in the inner ear.Prog Neurobiol. 1998 Feb;54(2):127-48. doi: 10.1016/s0301-0082(97)00054-3. Prog Neurobiol. 1998. PMID: 9481795 Review.
Cited by
-
Hair cell toxicology: With the help of a little fish.Front Cell Dev Biol. 2022 Dec 13;10:1085225. doi: 10.3389/fcell.2022.1085225. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 36582469 Free PMC article. Review.
-
Physiological responses of mechanosensory systems in the head of larval zebrafish (Danio rerio).Front Robot AI. 2023 Jul 31;10:1212626. doi: 10.3389/frobt.2023.1212626. eCollection 2023. Front Robot AI. 2023. PMID: 37583713 Free PMC article.
-
A Novel Functional Method of Protector Screening for Zebrafish Lateral Line Hair Cells via the Acoustic Escape Response.Neurosci Bull. 2025 Sep;41(9):1537-1552. doi: 10.1007/s12264-025-01406-3. Epub 2025 May 6. Neurosci Bull. 2025. PMID: 40329138
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous