

A photosynthesis operon in the chloroplast genome drives speciation in evening primroses
- PMID: 34048579
- PMCID: PMC8408503
- DOI: 10.1093/plcell/koab155
A photosynthesis operon in the chloroplast genome drives speciation in evening primroses
Erratum in
-
Erratum.Plant Cell. 2022 Jan 20;34(1):700. doi: 10.1093/plcell/koab233. Plant Cell. 2022. PMID: 34689204 Free PMC article. No abstract available.
Abstract
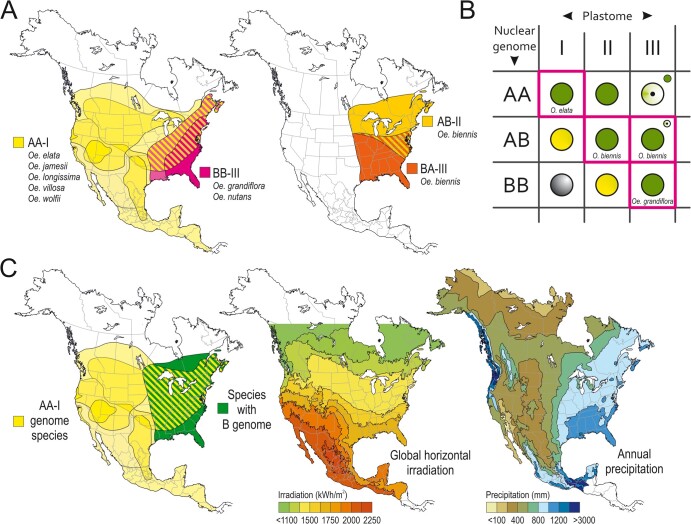
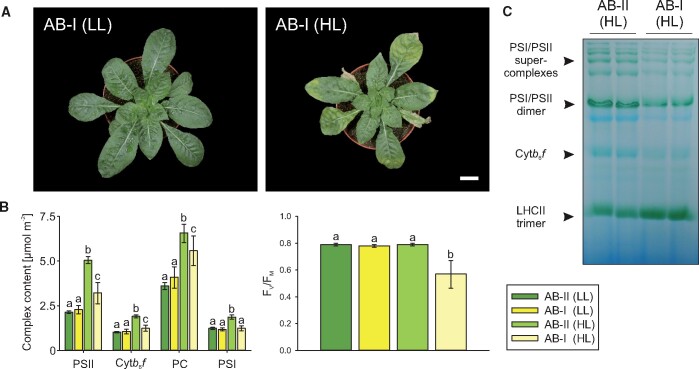
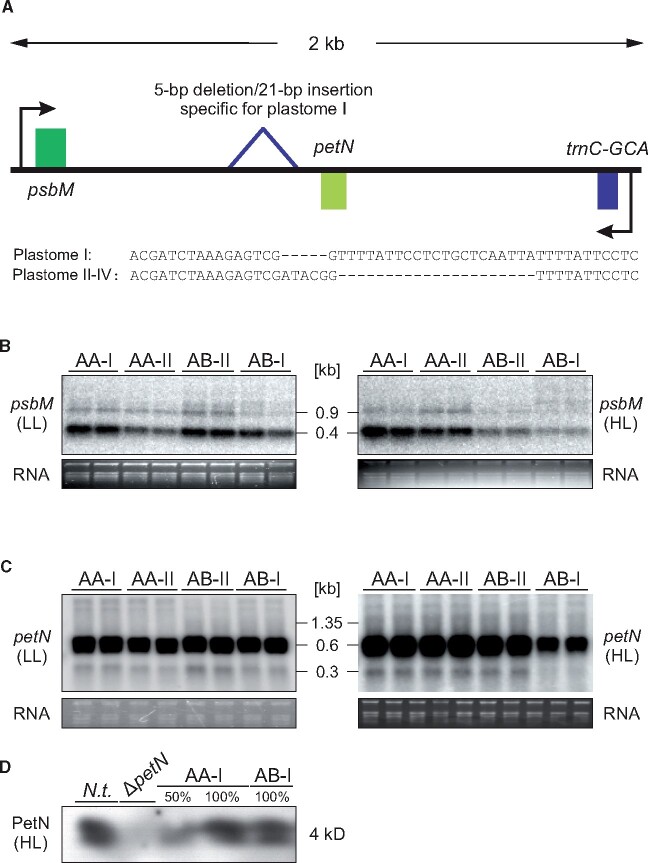
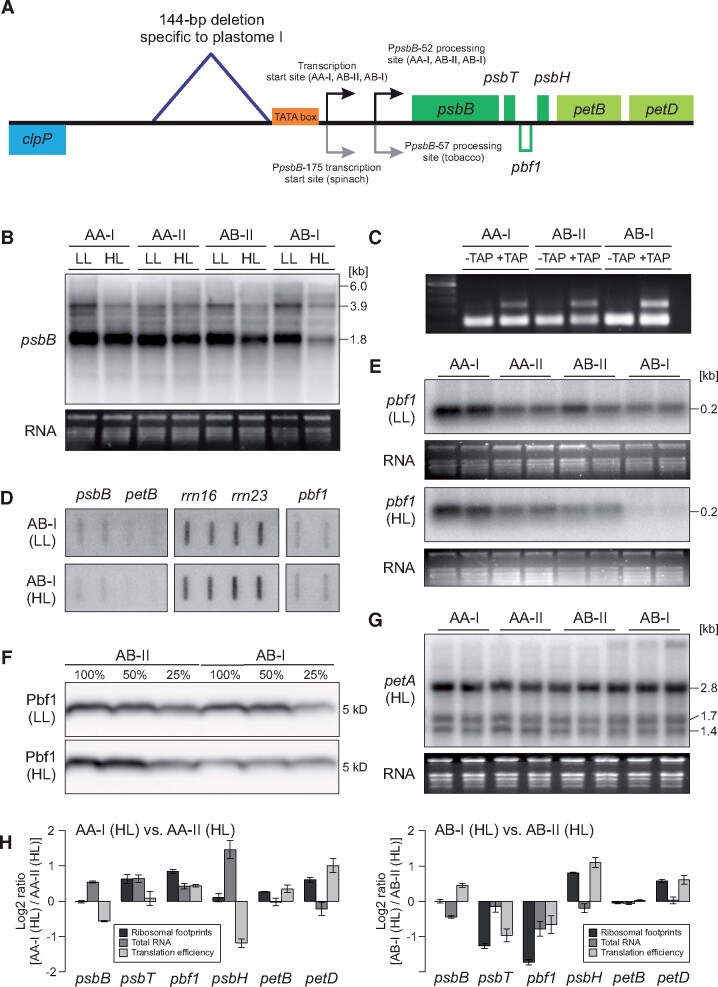
Genetic incompatibility between the cytoplasm and the nucleus is thought to be a major factor in species formation, but mechanistic understanding of this process is poor. In evening primroses (Oenothera spp.), a model plant for organelle genetics and population biology, hybrid offspring regularly display chloroplast-nuclear incompatibility. This usually manifests in bleached plants, more rarely in hybrid sterility or embryonic lethality. Hence, most of these incompatibilities affect photosynthetic capability, a trait that is under selection in changing environments. Here we show that light-dependent misregulation of the plastid psbB operon, which encodes core subunits of photosystem II and the cytochrome b6f complex, can lead to hybrid incompatibility, and this ultimately drives speciation. This misregulation causes an impaired light acclimation response in incompatible plants. Moreover, as a result of their different chloroplast genotypes, the parental lines differ in photosynthesis performance upon exposure to different light conditions. Significantly, the incompatible chloroplast genome is naturally found in xeric habitats with high light intensities, whereas the compatible one is limited to mesic habitats. Consequently, our data raise the possibility that the hybridization barrier evolved as a result of adaptation to specific climatic conditions.
© The Author(s) 2021. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Figures

References
-
- Arntz MA, Delph LF (2001) Pattern and process: evidence for the evolution of photosynthetic traits in natural populations. Oecologia 127: 455–467 - PubMed
-
- Barnard-Kubow KB, So N, Galloway LF (2016) Cytonuclear incompatibility contributes to the early stages of speciation. Evolution 70: 2752–2766 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
