

Notch-Wnt signal crosstalk regulates proliferation and differentiation of osteoprogenitor cells during intramembranous bone healing
- PMID: 34050174
- PMCID: PMC8163848
- DOI: 10.1038/s41536-021-00139-x
Notch-Wnt signal crosstalk regulates proliferation and differentiation of osteoprogenitor cells during intramembranous bone healing
Abstract
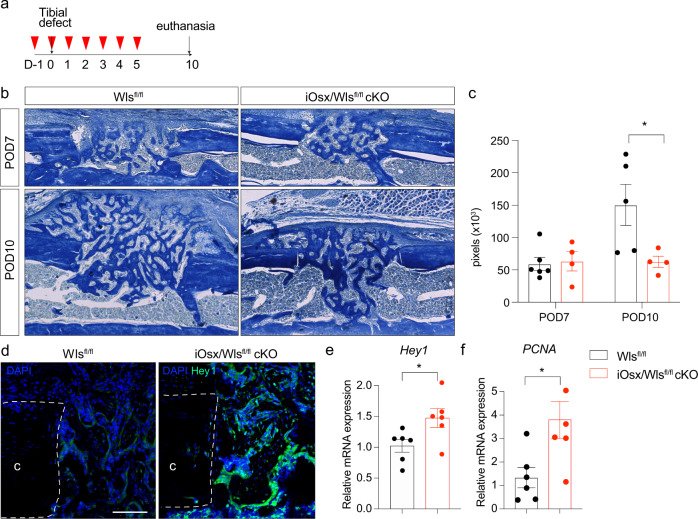
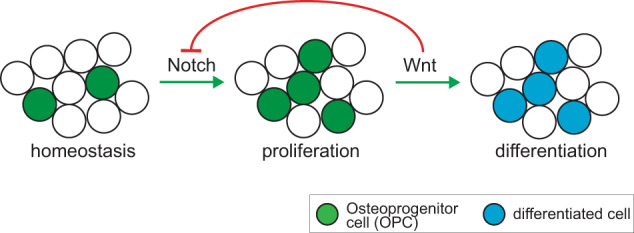
Adult bone regeneration is orchestrated by the precise actions of osteoprogenitor cells (OPCs). However, the mechanisms by which OPC proliferation and differentiation are linked and thereby regulated are yet to be defined. Here, we present evidence that during intramembranous bone formation OPC proliferation is controlled by Notch signaling, while differentiation is initiated by activation of canonical Wnt signaling. The temporospatial separation of Notch and Wnt signal activation during the early stages of bone regeneration suggests crosstalk between the two pathways. In vitro and in vivo manipulation of the two essential pathways demonstrate that Wnt activation leads to initiation of osteogenic differentiation and at the same time inhibits Notch signaling, which results in termination of the proliferative phase. Here, we establish canonical Wnt signaling as a key regulator that facilitates the crosstalk between OPC proliferation and differentiation during intramembranous, primary bone healing.
Conflict of interest statement
The authors declare no competing interests.
Figures
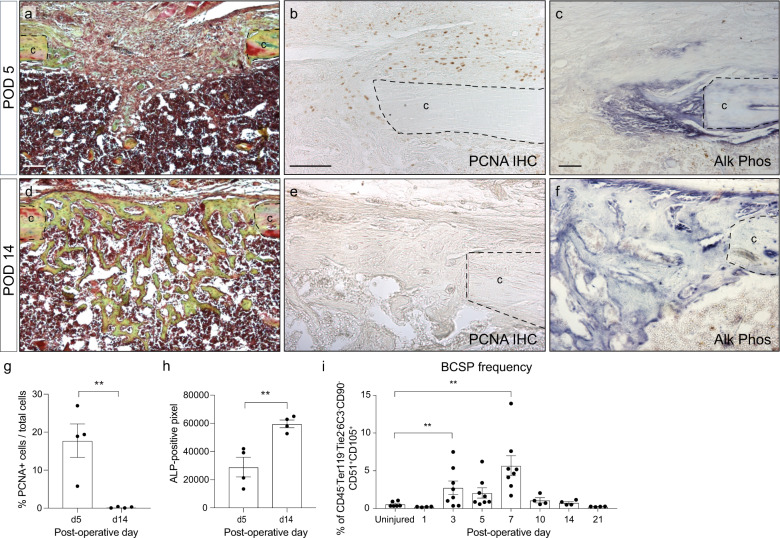
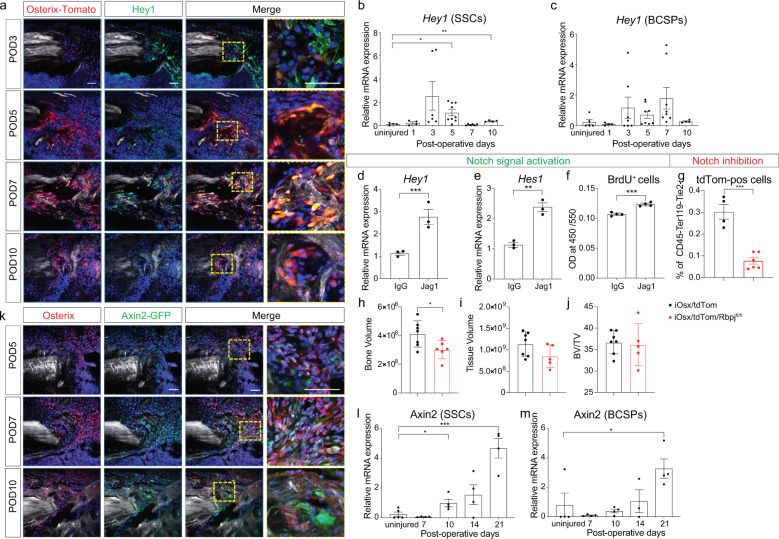
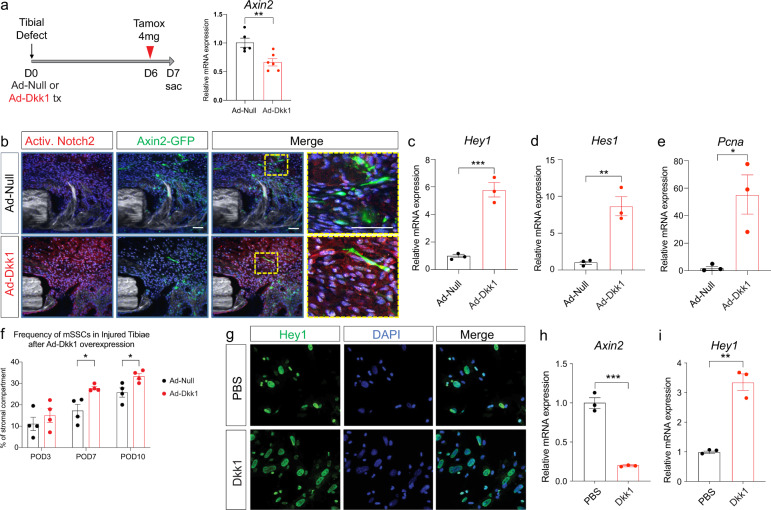
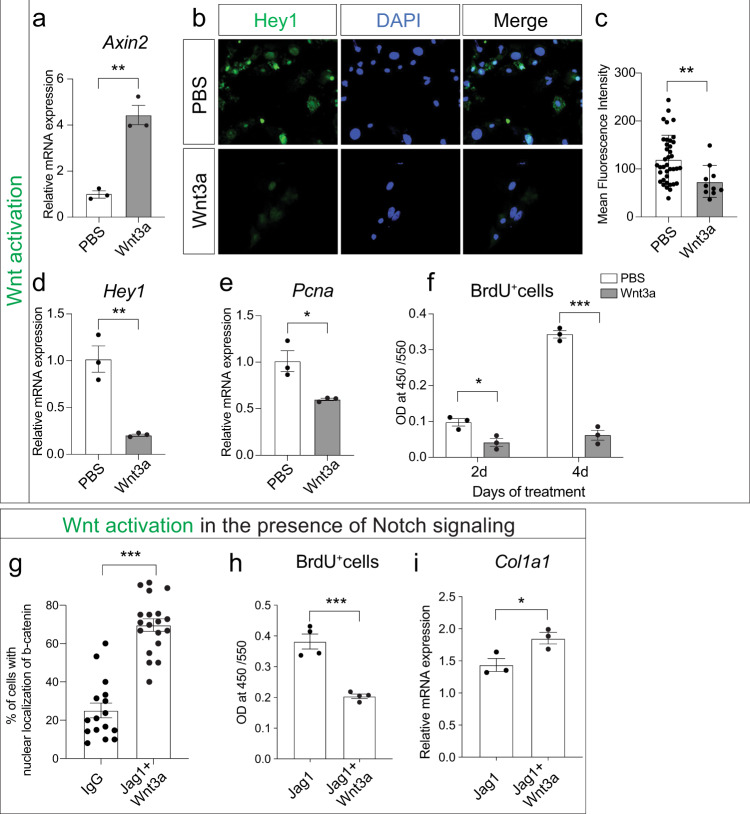
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
